
MOLECULAR AND CELLULAR BIOLOGY, Oct. 1996, p. 5821–5829 Vol. 16, No. 10
0270-7306/96/$04.0010
Copyright q 1996, American Society for Microbiology
DNA Length Is a Critical Parameter for Eukaryotic
Transcription In Vivo
JOCELYN E. KREBS AND MARIETTA DUNAWAY*
Department of Molecular and Cell Biology, University of California, Berkeley, Berkeley, California 94720-3202
Received 8 May 1996/Returned for modification 24 June 1996/Accepted 19 July 1996
The organization of eukaryotic chromosomes into topological domains has led to the assumption that DNA
topology and perhaps supercoiling are involved in eukaryotic nuclear processes. Xenopus oocytes provide a
model system for studying the role of DNA topology in transcription. Linear plasmid templates for RNA
polymerases (Pols) I and II are not transcribed in Xenopus oocytes, while circular templates are transcrip-
tionally active. Here we show that circularity is not required for transcription of Pol I or Pol II promoters if
the linear template is sufficiently long (>17 to 19 kb). The Xenopus rRNA (Pol I) promoter is active in central
positions on a long linear template but is not transcribed when located near an end. Because supercoils
generated by transcription could be retained by viscous drag against the long template, these results are
consistent with a supercoiling requirement for this promoter. Surprisingly, the herpes simplex virus thymidine
kinase (Pol II) promoter is active even 100 bp from the end of the long template, indicating that template length
fulfills a critical parameter for transcription that is not consistent with a supercoiling requirement. These
results show that DNA length has unrecognized importance for transcription in vivo.
Eukaryotic chromosomes are organized into topological do-
mains, so that a linear eukaryotic chromosome behaves like a
series of isolated closed loops (1, 5, 24). Because closed loops,
like circular DNAs, can be supercoiled, these topological do-
mains could potentially provide supercoiling for processes such
as transcription. However, despite extensive investigation, the
role of supercoiling and topological domains in eukaryotic
transcription is not clear. The classical approach for demon-
strating that a process requires supercoiling is to show that the
activity varies as a function of superhelical density. This ap-
proach has demonstrated that DNA supercoiling is required in
prokaryotes for initiation of DNA replication, site-specific re-
combination, and transcription of some genes (reviewed in
reference 14). This kind of analysis is possible in prokaryotes
because of the specificities of the prokaryotic topoisomerases.
In prokaryotes, DNA gyrase introduces negative supercoils or
relaxes positive supercoils and topoisomerase I relaxes only
negative supercoils; thus, the entire genome is more negatively
supercoiled in a topoisomerase I mutant. In contrast, eukary-
otic topoisomerases relax positive and negative supercoils
equally well. An additional complicating factor is that when
supercoiled plasmids are introduced into cells, they are imme-
diately relaxed by endogenous topoisomerases and subse-
quently re-supercoiled by packaging into nucleosomes. One
approach to overcoming these difficulties is to study the de-
pendence of transcription on supercoiling in vitro. Some eu-
karyotic genes are transcribed more efficiently in cell extracts if
they are supercoiled (13, 20, 30), although the presence of
topoisomerases in the extracts compromises the conclusion
that supercoiling facilitates transcription. Furthermore, exper-
iments show that the basal transcription factors IIE and IIH
are not required in a minimal in vitro transcription reaction if
the transcription template is negatively supercoiled (11, 23).
However, in one in vitro study the transcriptional activity of the
Saccharomyces cerevisiae rRNA promoter varied as a function
of superhelical density in transcription extracts prepared from
top1 top2 mutants (29). This evidence strongly suggests that
supercoiling facilitates transcription of this promoter.
There have also been suggestive results in vivo for the par-
ticipation of supercoiling in eukaryotic transcription. Linear-
ized plasmid DNAs are poorly transcribed in mammalian tis-
sue culture cells compared with circular forms of the same
templates (34), and linear plasmid templates for RNA poly-
merase (Pol) I and II genes are not transcribed at all in Xeno-
pus laevis oocytes (12, 26). Even when circular templates in
oocytes are allowed to establish transcription and are subse-
quently linearized by injection of a restriction enzyme, tran-
scription is dramatically reduced or entirely abolished (12, 26).
Linear and circular templates are equally stable in oocytes, so
the failure of linear templates to be transcribed is not due to
degradation of injected templates. Thus, circular templates
fulfill a critical requirement for eukaryotic transcription in
vivo. Nevertheless, it is not clear whether the requirement for
circular templates reflects a supercoiling requirement or
whether circularity per se is somehow required for transcrip-
tion. For example, free DNA ends in linear templates could be
inhibitory, perhaps because they provide an entry site for in-
hibitory proteins or initiate cooperative binding of an inhibitor.
Indeed, capping free DNA ends has previously been shown to
increase transcription of linear templates in transient transfec-
tion assays (6). To show a requirement for DNA supercoiling
in vivo therefore requires that we address the issue of whether
the ends of linear DNA are inhibitory and that we distinguish
a requirement for template circularity from a requirement for
DNA supercoiling.
One way to separate circularity from supercoiling is to gen-
erate supercoiled domains on a linear template. Transcription
itself generates positive supercoils ahead of the Pol and nega-
tive supercoils behind it if two critical requirements are ful-
filled (18). First, the RNA Pol must be restricted from rotating
freely around the DNA helix. Second, the supercoils that are
generated must not diffuse off the ends; this requirement can
be fulfilled by tethering the DNA to restrict its free rotation.
Transcription-driven supercoiling clearly occurs both in vitro
and in vivo, but the effect of transcription-driven supercoiling
on other cellular processes in vivo is uncertain (4, 10, 29, 31,
* Corresponding author. Phone: (510) 643-6208. Fax: (510) 642-
5821

37). Further, the importance of transcript length, DNA- and
RNA-binding proteins, chromosomal context, the competing
relaxation of supercoils by topoisomerases, and other aspects
of the nuclear environment in restricting rotation of RNA Pol
and in preventing diffusion of supercoils in vivo is not known.
We have previously separated template circularity from su-
percoiling by generating transcription-driven supercoils on a
linear template in the Xenopus oocyte (7). While endogenous
Xenopus Pol I or II cannot normally transcribe linear plasmid
templates, T7 RNA Pol is able to do so when injected into
oocytes. We showed that the Xenopus rRNA promoter can be
activated on a linear template by transcription from a divergent
upstream T7 RNA promoter. This and additional experiments
from that study suggest that transcription-driven supercoiling
can provide the minimum topological requirement for tran-
scription for this promoter (7). These results further suggest
that even a short linear DNA with no known tethers can, at
least transiently, retain supercoils at sufficient density to acti-
vate a promoter.
In this study we investigate the importance of template to-
pology and template length on transcription. We show that we
cannot activate transcription on a linear plasmid by competing
for possible inhibitory end-binding factors with DNA frag-
ments nor by blocking the ends of linear plasmids with DNA
hairpins. Most importantly, we show that both Pol I and Pol II
promoters are transcribed in long linear templates provided by
phage l cloning vectors. We have varied the positions of the
reporter genes within the l templates by circular permutation
in order to determine whether DNA flanking the promoters
was required. The rRNA promoter is transcribed only when it
is located far from the ends of the long linear template, sug-
gesting that the role of DNA length for this promoter is to
FIG. 1. Construction of hairpin-ended templates. (A) Diagram of method used to construct hairpin-ended templates. Hairpin-ended templates were made by
cutting plasmids with a restriction enzyme and then ligating them to hairpin-forming oligonucleotides. This is described in detail in Materials and Methods. (B)
Ethidium bromide-stained alkaline agarose gels of the hairpin-ended templates. The third lane of each gel shows the hairpin constructs immediately after the ligation
step. The fourth lane of each gel shows the result of exonuclease III digestion of the samples in the third lanes.
5822 KREBS AND DUNAWAY MOL.CELL.BIOL.
prevent or retard diffusion of transcription-driven supercoils.
However, the thymidine kinase (TK) promoter is active
whether it is centrally located or near an end, indicating that
the essential function of DNA length for this promoter is not,
or is not solely, to maintain supercoiling. Instead, DNA length
serves some other essential role in transcription. We discuss
several possibilities for the function of DNA length in tran-
scription, including the idea that DNA length overcomes a
kinetic barrier to assembly of the transcription complex by
facilitated diffusion of transcription factors to the promoter.
MATERIALS AND METHODS
Template construction. To make the l templates, the 760-bp rRNA minigene,
c40 (16), was cloned into the EcoRI site of lgt11. The 2-kb herpes simplex virus
TK gene (19) was cloned into the SalI and XbaI sites of lgt22A. To make the
multimerized plasmid template, two different plasmids containing the TK gene
(pBR322 and pBluescript) were linearized with the restriction enzyme XbaI and
then ligated to form concatamers. Ligation products were separated by agarose
gel electrophoresis, and high-molecular-weight products were isolated from the
gel. These gel-purified products were then used to transform DH5a cells. The
multimer plasmid was linearized with a restriction enzyme (XhoI) that cut in only
one of the two plasmids.
Hairpin-ended templates were made by cutting plasmids containing either the
TK gene or the rRNA minigene with the restriction enzyme XbaI and then
ligating the linearized plasmids to hairpin-forming oligonucleotides (59-CTAGC
GCTGCAGACTTTTTGTCTGCAGCG-39). The ligated samples were then di-
gested with exonuclease III to remove nicked and unligated material. Hairpin-
ended templates were analyzed on agarose gels under alkaline conditions (0.05
M NaOH, 1 mM EDTA), in which hairpin-ended templates run with the mobility
of single-stranded dimer circles. Nicked hairpin templates run as linear dimers
and are therefore separable on alkaline gels. All detectable nicked or unligated
material was digested by exonuclease III.
Analysis of oocyte injection experiments. All templates were microinjected
into the nuclei of X. laevis oocytes in a volume of 20 nl; 0.5 ng of linear and
circular plasmid templates and reference templates was injected unless otherwise
specified. Long linear templates were injected at approximately 5 ng in order to
achieve an equimolar ratio of experimental reporter gene to reference. rRNA
templates were coinjected with 50 mgofa-amanitin per ml. Otherwise, injections
and harvesting of oocyte nucleic acids were performed as described elsewhere
(16).
Two assays were employed to detect transcription from injected templates.
Transcription from the TK gene and its corresponding reference, ctk, was de-
tected by primer extension, as described previously (19). Transcription from the
rRNA minigene and reference was detected by S1 nuclease protection, as de-
scribed elsewhere (16). All transcription levels are normalized to the level of
transcription from the circular plasmid reference, which is given a value of one.
The normalized transcription levels also take into account any deviations from an
equal molar ratio in the injected templates (determined by Southern blotting [see
below]) and, in the case of the S1 protection assays, differences in the specific
activities of the S1 probes. Values for normalized transcription represent the
averages for at least three experiments, and the standard errors are given.
Transcription assays and Southern blots were quantitated with a Phosphor
Imager (Molecular Dynamics).
The molar ratios of the competing reporter genes and the stability of injected
templates were verified by Southern blotting of injected samples. All templates
used in this study were stable over the entire course of an injection experiment
(as long as 24 h at room temperature). In the cases in which the templates were
not injected at exact equal molar ratios, the differences in template levels were
taken into account in the final values for normalized transcription. The com-
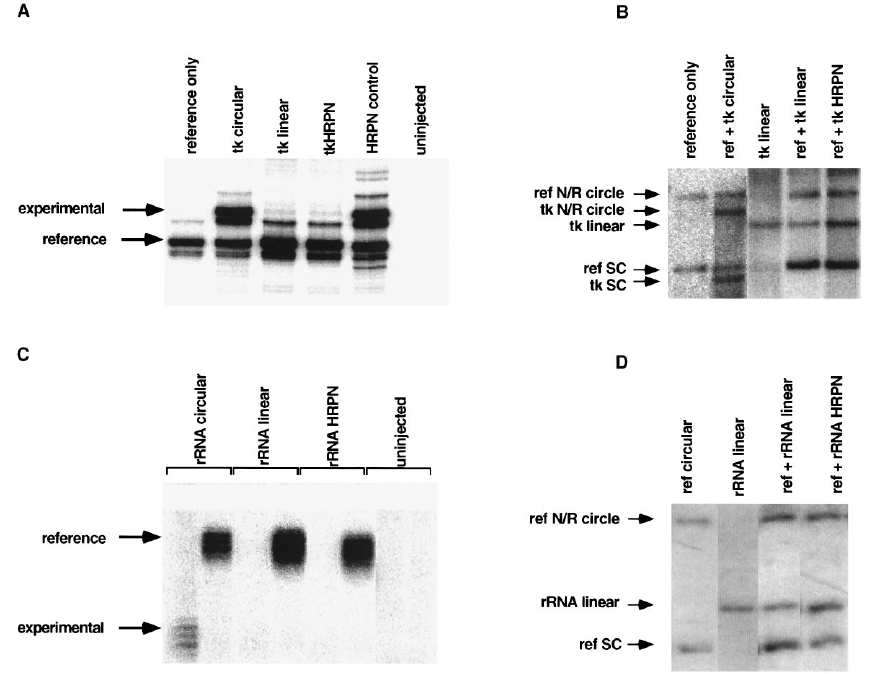
FIG. 2. Linear plasmid templates with hairpin ends are not transcribed. (A) Primer extension analysis of TK transcription by Pol II. The positions of the TK
(experimental) and the circular reference plasmid (reference) primer extension products are indicated by arrows. The band between the reference and experimental
signals is seen in the absence of experimental templates (see reference only lane) and is an artifact of the primer extension. (B) Southern blot of injected TK templates
after recovery from oocytes. (C) S1 nuclease analysis of rRNA transcription. The positions of the c40 (experimental) and the circular reference plasmid (reference)
S1-protected signals are indicated by arrows. (D) Southern blot of injected rRNA templates after recovery from oocytes. HRPN, hairpin; N/R, nicked/relaxed; SC, supercoiled.
VOL. 16, 1996 DNA TOPOLOGY AND LENGTH EFFECTS ON TRANSCRIPTION 5823

pleteness of digestion of l templates was determined by digesting the injected l
DNAs with HindIII or PvuI to produce multiple small fragments. These digested
samples were then Southern blotted and probed with HindIII or PvuI fragments
that contain the sites for the enzymes used for circular permutation or deletion
of l templates. Full-size HindIII or PvuI fragments would be seen only if diges-
tion by the first enzyme(s) was incomplete. Injection samples in which these
full-size fragments were detectable were discarded.
RESULTS
Free DNA ends do not inhibit transcription. We first tested
the possibility that linear templates are not transcribed because
of the presence of free DNA ends in these templates. Free
DNA ends could inhibit transcription because of binding or
entry of proteins at the ends. To test this possibility, we blocked
the free ends of linear templates by preparing linear plasmid
templates with hairpin ends. The construction of these tem-
plates is shown in Fig. 1. Plasmids were cut with a restriction
enzyme (XbaI) and then ligated to self-complementary oligo-
nucleotides. These oligonucleotides form hairpin structures
with a 59 single-strand extension that is complementary to the
restriction site. After ligation, the DNA is treated with exonu-
clease III to degrade any unligated or nicked templates (Fig.
1A; see Materials and Methods). The constructs were analyzed
on denaturing alkaline agarose gels, as shown in Fig. 1B. With
these gels it is possible to separate both monomer- and dimer-
length circles and linear DNAs. Dimer-length circles corre-
spond to hairpin-ended templates. Dimer-length linear DNAs
correspond to nicked hairpin templates or single-hairpin tem-
plates. These templates are sensitive to exonuclease III diges-
tion (Fig. 1B, 1EXO III). The identities of the bands on the
alkaline gels were confirmed by end labelling the hairpin oli-
gonucleotides with
32
P and subsequently exposing the alkaline
gels to film (data not shown).
In this and all other experiments reported here we have
tested two different transcription templates: the herpes simplex
virus TK gene, transcribed by RNA Pol II, and the X. laevis 40S
rRNA gene (rRNA) transcribed by RNA Pol I. In all cases, the
molar ratios and stability of injected templates were verified by
Southern blotting (Fig. 2B and D and data not shown; see
Materials and Methods).
Hairpin-ended templates for each promoter were coinjected
with the appropriate circular reference template, and tran-
scription was assayed (Fig. 2A and C). Neither the 6-kb TK
template nor the 4-kb rRNA template is detectably transcribed
when linearized (Fig. 2A, tk linear; Fig. 2C, rRNA linear). The
limit of detection for the rRNA S1 assay is approximately 1%
of the signal seen for circular templates. The limit of detection
for the TK primer extension assay is at least 0.1% of the signal
seen for the circular reference. Blocking the ends of these
templates with hairpins does not allow them to be transcribed
(Fig. 2A, tk HRPN; Fig. 2C, rRNA HRPN). As a control, the
same plasmids were linearized, self-ligated to recircularize,
and digested with exonuclease III to mimic the end-blocking
procedure. These control templates were transcribed normally,
showing that this procedure itself does not render templates
FIG. 3. Free DNA ends do not inhibit transcription. (A) Schematic of ex-
periment. Linear transcription template was coinjected with a circular reference
plasmid and various amounts of excess linear fragments (with EcoRI or SalI
termini). (B) Primer extension analysis of TK transcription with (tk linear 1 free
ends) and without (tk linear) excess free DNA ends. The triangle indicates
increasing molar excess of free ends of 2-fold, 20-fold, and 200-fold (left to
right).
FIG. 4. TK and rRNA promoters in linear l templates are transcribed. (A)
Templates used in the injection assay. The reporter genes used in previous
experiments were cloned into l vectors to create 45-kb linear templates. l
templates were injected at a molar ratio equal to that of the circular reference
plasmids. (B) Primer extension analysis of TK transcription. (C) S1 nuclease
analysis of rRNA transcription.
5824 KREBS AND DUNAWAY MOL.CELL.BIOL.

inactive (Fig. 2A, HRPN control). Southern blots of the in-
jected templates are also shown (TK templates, Fig. 2B; rRNA
templates, Fig. 2D). The hairpin-ended templates were still
resistant to exonuclease III digestion after reisolation from the
oocytes (data not shown).
We next attempted to compete for inhibitory end-binding
factors by coinjecting a linear template with increasing concen-
trations of free ends. No transcription was detected from linear
TK templates in the presence of a 2-, 20-, or 200-fold molar
excess of free ends (Fig. 3). The linear rRNA template also was
FIG. 5. Transcription of circular permutations of l templates. (A) Diagram of the procedure and consequences of circular permutation of the l templates. Circular
permutations were made by ligating the cos sites of l templates to form concatemers and then digesting the concatemers with the restriction enzymes indicated.
Linearization of injected templates was determined by Southern blotting. (B) Summary of transcription of lTK circular permutations. The direction of TK transcription
(arrow) and length of flanking DNA in kilobases (numbers above templates) are indicated. Transcription of l templates was normalized to transcription of the circular
reference template. Values represent the averages from at least three experiments, and standard errors are given. (C) Summary of transcription of lrRNA circular
permutations.
VOL. 16, 1996 DNA TOPOLOGY AND LENGTH EFFECTS ON TRANSCRIPTION 5825

not activated by competing free ends (data not shown). Since
no transcription was detected even at the highest levels of
competitor free ends and since hairpin-ended linear plasmids
are not transcriptionally active, it is unlikely that the failure of
short linear templates to be transcribed is due to the binding of
inhibitory factors to their free ends.
Long linear templates are transcriptionally active. We next
tested whether template length was a factor in transcription of
linear templates. We reasoned that long DNA arms flanking a
transcription template might act as a barrier to diffusion of
supercoils due to friction against the mass of DNA and pro-
teins. If this were the case, then negative supercoils generated
behind a Pol during transcription from a promoter in a long
template might be sustained long enough to facilitate further
initiation events.
Long linear templates were constructed by cloning either the
rRNA or the TK reporter gene into phage l vectors (Fig. 4A).
These templates are approximately 45 kb in length, and the
reporter genes are centrally located within the vector DNA
when linearized at the cos sites. The l templates were coin-
jected with an equimolar amount of circular reference plasmid,
and transcription was assayed (Fig. 4B and C). In dramatic
contrast to linear plasmids, both the rRNA and TK genes are
transcribed on long linear templates (Fig. 4B, lTK; Fig. 4C,
lrRNA).
The position of the rRNA promoter within a linear template
is critical for transcription. If transcription on the long linear
templates resulted from the formation of localized domains of
supercoiling due to a diffusion barrier provided by long DNA
arms, then transcription of a promoter in a long linear tem-
plate would be expected to depend on its position within the
template. Specifically, a promoter located near the end of a
template will not be transcribed as efficiently, since supercoils
diffuse rapidly off a short linear stretch of DNA. We tested the
importance of promoter position within the l templates by
making circular permutations of the l templates (Fig. 5A). The
cos ends of lTK or lrRNA were ligated in vitro to form a
mixture of concatemers and circular templates. Aliquots of
these mixtures were each digested with specific restriction en-
donucleases to create linear templates in which the position of
the reporter gene varies with respect to the ends of the tem-
plates. This circular permutation not only allowed us to test the
position dependence of transcription in these templates but
also allowed us to eliminate the possibility that the transcripts
detected from the l templates in Fig. 3 arose from templates in
which the l cos sites had annealed in vivo. The cos ends of the
l templates have much longer overhangs (12 bp) compared to
those generated by restriction enzyme cleavage of plasmid
DNAs. In the circularly permuted templates, the cos sites are
ligated and located internally, while the template ends are
generated by restriction enzymes like the linear plasmid tem-
plates. Permuted lrRNA templates were coinjected with cir-
cular reference plasmids, and transcription was assayed and
quantitated as described above (Fig. 5B). Templates in which
the rRNA promoter is still flanked with long DNA arms but
that have restriction enzyme overhangs instead of long com-
plementary ends from the cos sites are transcribed with effi-
ciency equal to that of the original l templates (Fig. 5B, cos
versus NheI). Therefore, the difference between transcription
on l templates and linear plasmid templates is not due to
FIG. 6. Transcription of shortened l templates. Shortened templates were made by digesting l templates with the restriction enzymes indicated, and the resultant
mixture of DNA fragments was coinjected with the reference plasmid into oocytes. (A) Summary of transcription of shortened lTK templates. (B) Summary of
transcription of shortened lrRNA templates. The direction of TK and rRNA transcription (arrows), the length of flanking DNA in kilobases (numbers above
templates), and total template length are indicated.
5826 KREBS AND DUNAWAY MOL.CELL.BIOL.

reannealing of the ends. We have also prevented annealing of
cos sites directly by filling in the cos overhangs with Klenow
Pol, and we see that these templates are transcribed to the
same extent as those shown in Fig. 4 (data not shown).
The position of the rRNA promoter was critical for tran-
scription in the long template, however. This promoter is not
transcribed when located within 100 bp of the end of the
template (Fig. 5B, KpnI and Asp718), regardless of whether the
end has a 39 or 59 overhang. This observation is consistent with
our original idea that long linear templates might allow do-
mains of supercoiling to accumulate and that this makes tran-
scription possible on a linear template.
Transcription of the TK promoter is independent of posi-
tion. To our surprise, TK and rRNA promoters do not have the
same dependence on position within the l template. In con-
trast to the rRNA promoter, moving the TK promoter close to
the end of the l template had only modest effects on transcrip-
tion. Transcription is only decreased about twofold in TK tem-
plates in which the TK gene is between 100 bp and 1 kb from
an end (Fig. 5C, SalI, ScaI, and XbaI). Transcription was ob-
served whether the 59 or 39 end of the gene was very close to
an end, suggesting that transcription from this promoter is not
dependent on supercoiling. DNA length is apparently provid-
ing some other critical function for this promoter.
A minimum length is required for a linear template to be
transcriptionally active. Since 45-kb linear l templates are
transcribed but 6-kb linear plasmids are not, we tested tem-
plates of intermediate sizes for transcriptional activity. The ltk
and lrRNA templates were digested with restriction enzymes
to create shortened templates, and the mixture of DNA frag-
ments was injected into oocytes. The shortened templates and
a summary of the transcription data are shown in Fig. 6. Both
the TK and rRNA promoters are active in templates as small
as 20 kb (Fig. 6A, XhoI, HindIII, and StuI-XhoI; Fig. 6B, BglII,
SnaBI-BglII, and StuI). However, neither promoter is active in
templates in the 10- to 14-kb range (Fig. 6A, ScaI-XhoI and
PvuI; Fig. 6B, PvuI).
Once a critical minimum length is reached, the level of
transcription increases with increasing length for the shortened
l templates (Fig. 7). We have not tested templates longer than
45 kb, but we note that transcription of these long linear
templates is still less than for a circular template. The mini-
mum length is similar for ltk and lrRNA, and the shape of the
curve is also similar. Since transcription does increase with
length in these templates, it seems that length alone can de-
termine efficiency of transcription of a template.
Linear plasmid multimers of sufficient size are transcribed.
To confirm that length rather than a sequence context was the
critical parameter for transcription of the l templates, we
constructed a long TK template by multimerizing plasmids that
are not transcribed as linear monomers (Fig. 8A). Two differ-
ent TK gene-containing plasmids (pBR322 and pBluescript)
were linearized with XbaI and ligated to form concatemers.
Neither of these TK gene-containing plasmids is transcribed as
a linear monomer (TK gene in pBR322, Fig. 2; TK gene in
pBluescript, data not shown). The ligated material was trans-
formed into Escherichia coli, and multimers having only one
pBluescript vector were selected. Thus, the multimerized plas-
mid shown can be linearized by digestion with any restriction
enzyme having a site unique to pBluescript.
Transcription of the 17-kb multimer TK template is shown
in Fig. 8B. The linear plasmid multimer is transcribed at ap-
FIG. 7. Transcription increases with increasing length. Plot of transcription level versus template length for lTK and lrRNA templates. Units of transcription are
arbitrary. The plot includes data from circular permutations and shortened templates.
VOL. 16, 1996 DNA TOPOLOGY AND LENGTH EFFECTS ON TRANSCRIPTION 5827

proximately one-third of the level of the circular reference
plasmid (normalized with respect to the copy number of TK
genes in the construct). This level of transcription is similar to
that of the long TK template provided by the l vector. This
indicates that length alone provides an essential function for
transcription of linear templates.
DISCUSSION
We have shown that long linear templates are transcribed by
both Pol I and Pol II with an efficiency comparable to that of
circular plasmids. Transcription is not due to specific se-
quences in the l vector, because linear plasmid multimers of
sufficient length are transcribed. Thus, a linear plasmid tem-
plate that is transcriptionally inactive can be activated simply
by increasing its length. The minimum length required for
transcription of either RNA Pol I or Pol II templates is be-
tween 14 and 17 kb, only a three- to fivefold increase over the
length of typical plasmids. A similar effect is observed in tissue
culture transfection experiments, in which circular templates
are preferred to linear templates when the templates are at low
concentrations, but at high DNA concentrations, linear tem-
plates are transcribed. In these cases, the high DNA concen-
tration leads to concatemerization of the linear plasmids (25,
33, 34). Because there is no end-joining activity in Xenopus
oocytes, short linear templates do not form concatemers even
when they are injected at high concentrations in our assays.
It is clear from the experiments reported here that DNA
length facilitates transcription of RNA Pol I and II genes in
different ways. Transcription of the Xenopus rRNA promoter is
dependent on the position of the promoter with respect to the
ends of the DNA, consistent with the idea that a topological
domain can be created on a large linear template. The viscous
drag against a large DNA molecule is apparently sufficient to
prevent transcription-generated supercoils from diffusing rap-
idly off the end of the DNA, so that DNA length creates a
topological domain. These results, especially when taken with
previous data showing that transcription-driven supercoiling
can activate the rRNA promoter (7), suggest that this pro-
moter requires supercoiling. The two studies taken together
are the best evidence to date that supercoiling is required for
any function in vivo in a eukaryote.
In contrast to rRNA transcription, the transcription of the
TK promoter is independent of its position within the l tem-
plate. Templates in which either the 59 or 39 end of the TK
gene is near the end are transcribed with efficiency similar to
the efficiency of those in which the TK promoter is centrally
located within the l DNA. These observations suggest that
long DNA serves different functions for the two genes tested
and separates transcription of the two genes mechanistically.
This distinction is further supported by the experiments using
T7 transcription to activate eukaryotic promoters. The TK
promoter could not be activated by transcription from a diver-
gent T7 promoter (6a).
If the results with TK are not consistent with a supercoiling
requirement, then what is the advantage to having a long
template for transcription? A classic problem in molecular
biology is how a sequence-specific DNA-binding protein finds
its target sequence within the vast excess of nonspecific DNA
(17, 32). Given the relative affinities of sequence-specific
DNA-binding proteins for their target sequences and for non-
specific DNA, these proteins would never find their targets if
the nonspecific DNA acted only as a competitor. Our experi-
ments show that the additional nonspecific DNA present in cis
on the long templates facilitates transcription and therefore
cannot be considered a competitor in the traditional sense.
There is previous evidence that DNA length can facilitate a
process. The rate at which site-specific DNA-binding proteins
find their sites is increased (8, 15, 35, 36). For example, the
association of lac repressor with an operator sequence is ap-
proximately 10-fold faster in a l template than in an oligonu-
cleotide duplex (2, 27, 28). This rate is about 1,000 times faster
than one would expect for a diffusion-controlled macromolec-
ular association. The increased rate of association on long
DNA is proposed to occur by a process of facilitated diffusion
in which DNA-binding proteins associate with nonspecific
DNA and either slide or hop along the DNA contour or are
directly transferred from one segment of DNA to another (3,
32). This reduces a three-dimensional search to a one- or
two-dimensional search. Thus, the activation of the TK pro-
moter on long templates fulfills one criterion discussed by von
FIG. 8. Linear plasmid multimers are transcribed. (A) Construction of the
TK multimer template. Two different plasmids containing the TK gene (pBR322
and pBluescript SK) were linearized with the restriction enzyme XbaI and then
ligated to form concatemers. Ligation products were separated by agarose gel
electrophoresis, and high-molecular-weight products were isolated from the gel.
These gel-purified products were then used to transform E. coli. The multimer
plasmid was linearized with a restriction enzyme (XhoI) that cuts in pBluescript
only. (B) Primer extension analysis of TK transcription of multimerized tem-
plates.
5828 KREBS AND DUNAWAY MOL.CELL.BIOL.
Hippel and Berg as indicative of a process in which facilitated
diffusion is important: the rate of the process is increased when
template length is increased (32). Although our experiments
do not address which step or steps in the transcription process
are rate limiting in these different templates, it is possible that
the increased rate at which sequence-specific transcriptional
activators find their targets facilitates assembly of the tran-
scription complex, perhaps by providing a surface for nucle-
ation of the complex. There are other possible roles for a long
template, such as attachment to the nuclear architecture or
targeting to a nuclear compartment rich in transcription fac-
tors. Attachment, for example, could create DNA loops that
mimic the topological domains of the chromosome. In either
case, our results show that such attachment or targeting must
be triggered solely by DNA length, not specific sequences.
There are other examples of the importance of DNA length
in cellular processes. DNA length is important for chromo-
some segregation and nuclear assembly (9, 21, 22), though the
role of DNA length is not understood for either process. It is
clear that the dynamics of long DNAs play a critical role in
many nuclear activities, and studies of these dynamics will lead
to greater understanding not only of these processes but of the
structure and function of the chromosome itself.
ACKNOWLEDGMENTS
We thank N. Cozzarelli, M. Botchan, R. Harland, L. Zechiedrich, S.
Uptain, and C. Robinett for critical reading of the manuscript.
This work was supported by a grant to M.D. from the National
Institute of General Medical Sciences.
REFERENCES
1. Benyajati, C., and A. Worcel. 1976. Isolation, characterization, and structure of
the folder interphase genome of Drosophila melanogaster. Cell 9:393–407.
2. Berg, O. G., and M. Ehrenberg. 1982. Association kinetics with coupled
three- and one-dimensional diffusion. Chain length dependence of the asso-
ciation rate of specific DNA sites. Biophys. Chem. 15:41–51.
3. Berg, O. G., R. B. Winter, and P. H. von Hippel. 1981. Diffusion-driven
mechanisms of protein translocation on nucleic acids. Models and theory.
Biochemistry 20:6929–6948.
4. Brill, S. J., and R. Sternglanz. 1988. Transcription-dependent DNA super-
coiling in yeast DNA topoisomerase mutants. Cell 54:403–411.
5. Cook, P. R., and I. A. Brazell. 1975. Supercoils in human DNA. J. Cell Sci.
19:261–279.
6. Courey, A. J. 1986. Structural and functional aspects of DNA supercoiling.
Ph.D. thesis. Harvard University, Cambridge, Mass.
6a.Dunaway, M., and E. Ostrander. Unpublished observations.
7. Dunaway, M., and E. A. Ostrander. 1993. Local domains of supercoiling
activate a eukaryotic promoter in vivo. Nature (London) 361:746–748.
8. Ehbrecht, H.-J., A. Pingoud, C. Urbanke, G. Maas, and C. Gualerzi. 1985.
Linear diffusion of restriction endonucleases on DNA. J. Biol. Chem. 260:
6160–6166.
9. Forbes, D. J., M. W. Kirschner, and J. W. Newport. 1983. Spontaneous
formation of nucleus-like structures around bacteriophage DNA microin-
jected into Xenopus eggs. Cell 34:13–23.
10. Giaever, G. N., and J. C. Wang. 1988. Supercoiling of intracellular DNA can
occur in eukaryotic cells. Cell 55:849–856.
11. Goodrich, J. A., and R. Tjian. 1994. Transcription factors IIE and IIH and
ATP hydrolysis direct promoter clearance by RNA polymerase II. Cell 77:
145–156.
12. Harland, R. M., H. Weintraub, and S. L. McKnight. 1983. Transcription of
DNA injected into Xenopus oocytes is influenced by template topology.
Nature (London) 301:38–43.
13. Hirose, S., and Y. Suzuki. 1988. In vitro transcription of eukaryotic genes is
affected differently by the degree of DNA supercoiling. Proc. Natl. Acad. Sci.
USA 85:718–722.
14. Kanaar, R., and N. R. Cozzarelli. 1992. Roles of supercoiled DNA structure
in DNA transactions. Curr. Opin. Struct. Biol. 2:369–379.
15. Khoury, A. M., H. J. Lee, M. Lillis, and P. Lu. 1990. Lac repressor-operator
interaction: DNA length dependence. Biochim. Biophys. Acta 1087:55–60.
16. Labhart, P. L., and R. H. Reeder. 1984. Enhancer-like properties of the
60-81 bp elements in the ribosomal gene spacer of Xenopus laevis. Cell
37:285–289.
17. Lin, S.-Y., and A. D. Riggs. 1975. The general affinity of lac repressor for E.
coli DNA: implications for gene regulation in procaryotes and eucaryotes.
Cell 4:107–111.
18. Liu, L. F., and J. C. Wang. 1987. Supercoiling of the DNA template during
transcription. Proc. Natl. Acad. Sci. USA 84:7024–7027.
19. McKnight, S. L., E. R. Gavis, and R. Kingsbury. 1981. Analysis of transcrip-
tional regulatory signals of the HSV thymidine kinase gene: identification of
an upstream control region. Cell 25:385–398.
20. Mizushima-Sugano, J., and R. G. Roeder. 1986. Cell-type-specific transcrip-
tion of an immunoglobulin kappa light chain gene in vitro. Proc. Natl. Acad.
Sci. USA 83:8511–8515.
21. Murray, A. W., N. P. Schultes, and J. W. Szostak. 1986. Chromosome length
controls mitotic segregation in yeast. Cell 45:529–536.
22. Murray, A. W., and J. W. Szostak. 1983. Construction of artificial chromo-
somes in yeast. Nature (London) 305:189–193.
23. Parvin, J. D., and P. A. Sharp. 1993. DNA topology and a minimal set of
basal factors for transcription by RNA polymerase II. Cell 73:533–540.
24. Paulson, J. R., and U. K. Laemmli. 1977. The structure of histone-depleted
metaphase chromosomes. Cell 12:817–828.
25. Perucho, M., D. Hanahan, and M. Wigler. 1980. Genetic and physical link-
age of exogenous sequences in transformed cells. Cell 22:309–317.
26. Pruitt, S. C., and R. H. Reeder. 1984. Effect of topological constraint on
transcription of ribosomal DNA in Xenopus oocytes: comparison of plasmid
and endogenous genes. J. Mol. Biol. 174:121–139.
27. Riggs, A. D., S. Bourgeois, and M. Cohn. 1970. Lac repressor-operator
interaction. Kinetic studies. J. Mol. Biol. 53:401–417.
28. Riggs, A. D., H. Suzuki, and S. Bourgeois. 1970. Lac repressor-operator
interaction. Equilibrium kinetics. J. Mol. Biol. 48:67–83.
29. Schultz, M. C., S. J. Brill, Q. Ju, R. Sternglanz, and R. H. Reeder. 1992.
Topoisomerases and yeast rRNA transcription: negative supercoiling stim-
ulates initiation and topoisomerase activity is required for elongation. Genes
Dev. 6:1332–1341.
30. Sive, H. L., N. Heintz, and R. G. Roeder. 1986. Multiple sequence elements
are required for maximal in vitro transcription of a human histone H2B gene.
Mol. Cell. Biol. 6:3329–3340.
31. Tsao, Y. P., H. Y. Wu, and L. F. Liu. 1989. Transcription-driven supercoiling
of DNA: direct biochemical evidence from in vitro studies. Cell 56:111–118.
32. von Hippel, P. H., and O. G. Berg. 1989. Facilitated target location in
biological systems. J. Biol. Chem. 264:675–678.
33. Weber, F., and W. Schaffner. 1985. SV40 enhancer increases RNA polymer-
ase density within the linked gene. Nature (London) 315:75–77.
34. Weintraub, H., P. F. Cheng, and K. Conrad. 1986. Expression of transfected
DNA depends on DNA topology. Cell 46:115–122.
35. Winter, R., and P. H. von Hippel. 1981. Diffusion-driven mechanisms of
protein translocation on nucleic acids. The Escherichia coli repressor-oper-
ator interaction: equilibrium measurements. Biochemistry 20:6948–6960.
36. Winter, R. B., O. G. Berg, and P. H. von Hippel. 1981. Diffusion-driven
mechanisms of protein translocation on nucleic acids. The Escherichia coli
lac repressor-operator interaction: kinetic measurements and conclusions.
Biochemistry 20:6961–6977.
37. Wu, H.-Y., S. Shyy, J. C. Wang, and L. F. Liu. 1988. Transcription generates
positively and negatively supercoiled domains in the template. Cell 53:433–
440.
VOL. 16, 1996 DNA TOPOLOGY AND LENGTH EFFECTS ON TRANSCRIPTION 5829
