
JOURNAL
OF
VIROLOGY,
JUlY
1993,
p.
4027-4036
0022-538X/93/074027-10$02.00/0
Copyright
©
1993,
American
Society
for
Microbiology
The
Two
Zinc
Fingers
in
the
Human
Immunodeficiency
Virus
Type
1
Nucleocapsid
Protein
Are
Not
Functionally
Equivalent
ROBERT
J.
GORELICK,`*
DONALD
J.
CHABOT,'
ALAN
REIN,2
LOUIS
E.
HENDERSON,'
AND
LARRY
0.
ARTHUR'
AIDS
Vaccine
Development
Program,
Program
Resources
Inc.,
DynCorp,
1
and
Laboratory
of
Molecular
Virology
and
Carcinogenesis,
Advanced
BioScience
Laboratories,
Inc.
-Basic
Research
Program,
2
National
Cancer
Institute-Frederick
Cancer
Research
and
Development
Center,
Frederick,
Maryland
21702-1201
Received
18
December
1992/Accepted
31
March
1993
The
highly
conserved
zinc
fingers
in
retroviral
nucleocapsid
(NC)
proteins
have
the
general
structure
Cys-(X)2-Cys-(X)4-His-(X)4-Cys.
Human
immunodeficiency
virus
type
1
(HIV-1)
contains
two
Zn2+
fingers,
and
mutants
were
constructed
in
which
the
native
sequence
of
each
Zn21
finger
was
maintained
but
their
positions
in
the
NC
protein
were
changed.
Mutants
had
either
two
first-finger
sequences
(pNC1/1),
two
second-finger
sequences
(pNC2/2),
or
reversed
first-
and
second-finger
sequences
(pNC2/1).
Cells
transfected
with
mutant
or
wild-type
clones
produced
similar
levels
of
Tat,
Gag,
Pol,
and
Env
proteins,
formed
syncytia,
and
shed
viruslike
particles
that
were
indistinguishable
by
electron
microscopy.
However,
the
pNC2/1
and
pNC2/2
mutants
were
inefficient
in
packaging
genomic
RNA
(less
than
15%
of
wild-type
levels),
whereas
the
pNCl/l
mutant
packaged
approximately
70%o
of
wild-type
levels
of
RNA.
No
infectious
virus
could
be
detected
with
either
the
pNC2/1
or
pNC2/2
mutants,
whereas
the
pNCl/l
mutant
appeared
to
sustain
a
low
level
of
replication
and
reverted
to
a
competent
wild-type-like
viral
species
after
a
2-
to
4-week
lag
period.
The
data
strongly
suggest
that
the
two
Zn2+
fingers
of
HIV-1
are
not
functionally
equivalent
and
that
the
first
Zn2+
finger
in
the
Gag
precursor
plays
a
more
prominent
role
in
RNA
selection
and
packaging.
The
data
also
indicate
that
both
Zn2+
fingers
in
the
mature
NC
protein
play
as
yet
unknown
roles
in
viral
assembly
or
the
early
stages
of
the
viral
infection
process.
All
retroviral
gag
genes
encode
a
Gag
precursor
polypro-
tein
that
functions
during
viral
assembly
to
form
the
inner
core
of
a
budding
virion.
The
Gag
precursor
selectively
packages
the
viral
RNA.
After
budding,
the
Gag
polyprotein
is
proteolytically
cleaved
into
structural
proteins
that
form
the
infectious
virus.
One
of
the
cleavage
products
is
a
small
basic
protein
that
binds
single-stranded
RNA
(13)
and
is
referred
to
as
the
nucleocapsid
(NC)
protein
(15).
All
retro-
viral
NC
proteins
contain
peptide
segments
consisting
of
14
amino
acid
residues
with
cysteine
and
histidine
residues
arranged
as
follows:
Cys-(X)2-Cys-(X)4-His-(X)4-Cys
(3,
5).
These
peptide
segments
chelate
Zn2+
ions
through
the
cysteine
sulfurs
and
the
histidine
imidazole
nitrogen
(4b,
17,
21,
25,
28).
Each
of
these
segments
forms
a
three-looped
structure
and
is
referred
to
as
a
zinc
finger
(3,
25).
Gag
precursors
and
NC
proteins
of
mammalian
type
C
retrovi-
ruses
(of
which
the
prototype
is
Moloney
murine
leukemia
virus)
contain
one
Zn2+
finger.
All
other
retroviruses
(except
the
spumaretroviruses,
which
do
not
contain
any
Zn
+
fingers
in
their
Gag
proteins,
according
to
known
nucleic
acid
sequences)
have
two
Zn2+
fingers
per
NC
protein,
including
avian
type
C
(prototype,
Rous
sarcoma
virus),
type
B
(prototype,
mouse
mammary
tumor
virus),
and
type
D
(prototype,
Mason-Pfizer
monkey
virus)
viruses,
human
T-lymphotropic
virus
types
I
and
II
(HTLV-I
and
HTLV-II,
respectively),
nonprimate
lentiviruses
(prototype,
equine
infectious
anemia
virus),
and
primate
lentiviruses
such
as
human
immunodeficiency
virus
types
1
and
2
(HIV-1
and
HIV-2,
respectively)
and
simian
immunodeficiency
virus.
*
Corresponding
author.
We
will
refer
to
these
as
two-fingered
NC
proteins
and
focus
attention
on
the
functions
of
the
individual
fingers
in
these
proteins.
Analysis
of
NC
mutants
with
altered
Zn2+-chelating
resi-
dues
(Cys
or
His)
has
shown
that
retroviruses
require
intact
Zn2+
fingers
for
specific
packaging
of
viral
RNA
(2,
6,
9,
10,
19).
Additionally,
retroviruses
with
two-fingered
NC
pro-
teins
require
both
fingers.
Therefore,
if
residues
are
mutated
such
that
either
finger
is
unable
to
chelate
Zn2+,
the
resulting
virus
is
noninfectious
and
deficient
in
packaged
viral
RNA
(2,
6,
10,
19).
Comparison
of
the
amino
acid
sequences
of
all
two-
fingered
retroviral
NC
proteins
reveals
that
the
first
finger
(from
the
N-terminal
end)
is
more
highly
conserved
than
the
second
finger
(20,
26).
The
first
fingers
of
all
two-fingered
NC
proteins
have
the
general
structure
Cys-Aro-Xb-CyS-Xc-Xd-
Xe-Gly-is-Xg-Xh-Xi-Car-Cys,
where
Aro
refers
to
an
aro-
matic
residue
(Trp,
Phe,
Tyr,
or
His)
and
Car
refers
to
a
residue
containing
a
carbonyl
group
(Asp,
Asn,
Glu,
or
Gln).
The
lentiviruses
retain
the
Aro,
Gly,
and
Car
residues
in
the
second
finger,
whereas
the
second
finger
of
all
other
retro-
viral
NC
proteins
have
substitutions
for
either
the
Aro
residue
or
the
Gly
residue
or
both.
Within
the
primate
lentivirus
subgroup
(HIV-1,
HIV-2,
and
simian
immunode-
ficiency
virus),
residues
at
positions
Xe
and
Xg
are
variable
in
both
fingers.
Residues
at
Xd,
Xh,
and
Xi
are
more
highly
conserved
in
the
first
finger
than
in
the
second
finger
(20).
These
observations
suggest
that
the
first
and
second
Zn2+
fingers
may
not
be
under
equal
evolutionary
selection
pres-
sures.
Therefore,
even
though
both
Zn2+
fingers
of
HIV-1
are
capable
of
binding
Zn2+
and
have
the
conserved
Aro,
4027
Vol.
67,
No.
7

4028
GORELICK
ET
AL.
Gly,
and
Car
residues,
they
still
may
not
be
functionally
equivalent.
To
test
this
hypothesis,
we
have
rearranged
the
Zn2+
fingers
of
HIV-1.
We
have
created
NC
mutants
in
which
all
of
the
conserved
ligand-binding
elements
are
intact
and
the
Zn2+
fingers
are
capable
of
coordinating
Zn2+.
Furthermore,
these
mutants
use
only
HIV-1
coded
sequences.
The
viral
coding
sequences
for
the
individual
Zn2+
fingers
have
been
duplicated
or
rearranged
as
described
below.
In
this
study
we
show
that
the
first
and
second
Zn2+
fingers
of
the
HIV-1
NC
protein
are
not
interchangeable
and
are
not
functionally
equivalent.
More
importantly,
we
provide
insights
into
pos-
sible
functions
of
the
individual
Zn2+
fingers
in
the
Gag
precursor
and
the
NC
proteins.
In
addition
to
genomic
RNA
packaging,
this
study
clearly
shows
that
Zn2+
fingers
in
the
Gag
precursor
and
NC
protein
are
also
involved
in
events
vital
to
early
infection
processes.
MATERIALS
AND
METHODS
Plasmids,
bacteria,
and
cell
lines.
The
pNLA-3
plasmid,
containing
an
infectious
proviral
clone
of
HIV-1
(1),
was
a
gift
of
Malcom
A.
Martin,
National
Institute
of
Allergy
and
Infectious
Diseases.
The
pBluescript
KS'
plasmid
used
for
mutagenesis
and
subcloning
was
from
Stratagene,
La
Jolla,
Calif.
DNA
transformations
were
performed
in
either
TG1
bacteria
(Amersham,
Arlington
Heights,
Ill.)
or
competent
DH10B
bacteria
(BRL/GIBCO,
Gaithersburg,
Md.).
DNA
plasmids
that
were
to
be
digested
with
Bcll
(a
methylation-
sensitive
restriction
endonuclease)
were
first
cultivated
in
Eschenchia
coli
GM2163
(New
England
BioLabs,
Inc.,
Beverly,
Mass.).
HeLa
cells
were
a
gift
of
George
N.
Pavlakis
and
Barbara
K.
Felber,
Advanced
BioScience
Laboratories,
Inc.-Basic
Research
Program,
National
Can-
cer
Institute
(NCI)-Frederick
Cancer
Research
and
Devel-
opment
Center
(FCRDC).
CD4-producing
HeLa
cells
con-
taining
a
long
terminal
repeat-3-galactosidase
construct
(HCLZ
cells)
were
constructed
by
and
obtained
from
David
Waters,
Program
Resources
Inc./DynCorp,
NCI-FCRDC
(31).
The
long
terminal
repeat-,-galactosidase
construct
was
used
to
detect
the
presence
of
HIV-1
Tat
protein
production.
HCLZ
cells
were
developed
for
1-galactosidase
expression
as
reported
previously
(16).
HeLa
and
HCLZ
cells
were
maintained
in
minimum
essential
medium
containing
Earle's
salts
and
10%
(vol/vol)
fetal
calf
serum
at
37°C
in
a
5%
CO2
atmosphere.
H9
cells
were
obtained
from
Robert
C.
Gallo,
NCI
(22),
and
maintained
in
RPMI
1640
medium
plus
5%
fetal
calf
serum,
10
mM
N-2-hydroxyethylpiperazine-N'-2-
ethanesulfonic
acid
(HEPES;
pH
7.3)
and
2
,g
of
Polybrene
per
ml
at
37°C
in
a
5%
CO2
atmosphere.
Mutagenesis.
Oligonucleotides
were
synthesized
by
the
Nucleic
Acid
and
Protein
Synthesis
Laboratory,
Program
Resources
Inc./DynCorp,
NCI-FCRDC,
on
a
Biosearch
model
8750
multiple-column
DNA
synthesizer.
Oligonucle-
otide-directed
mutagenesis
was
performed
by
using
the
Muta-Gene
Phagemid
In
Vitro
Mutagenesis
Kit,
Version
2
(Bio-Rad,
Richmond,
Calif.).
Because
of
the
homology
between
the
center
sections
of
the
oligonucleotides
used
for
oligonucleotide-directed
muta-
genesis
of
Zn2+
finger
coding
sequences
and
the
Zn2+
finger
sequences
that
are
already
present
in
proviral
clones,
we
subcloned
the
two
Zn2+
fingers
into
separate
plasmids
to
ensure
proper
annealing
of
the
mutagenic
oligonucleotides.
These
plasmids
were
mutagenized
to
exchange
the
first
and
second
Zn2+
finger
nucleotide
sequences.
Full-length
provi-
ruses
were
constructed
with
two
first-finger
sequences
(pNC1/1),
two
second-finger
sequences
(pNC2/2),
or
re-
versed
sequences
(pNC2/1).
As
described
below,
this
method
of
construction
does
not
generate
any
mutations
outside
the
Zn2+
finger
sequences.
Figure
1
is
a
diagram
depicting
the
construction
of
the
Zn2+
finger
rearrangement
mutants.
The
subclone,
desig-
nated
pRG1,
consists
of
a
4,278-bp
SpeI-SalI
fragment
from
pNL4-3
(pNC1/2)
that
was
cloned
into
the
corresponding
sites
of
pBluescript
KS'
as
described
previously
(10).
The
ApaI
site
in
the
polylinker
of
pRG1
was
eliminated
in
the
following
manner,
to
create
a
clone
with
a
unique
ApaI
site
between
the
sequences
that
code
for
the
two
Zn2+
fingers
(called
pDRO).
The
pRG1
plasmid
was
partially
digested
with
ApaI,
and
an
oligonucleotide
with
the
sequence
5'-TGG
ATC
CAT
GGA
TCC
AGG
CC-3'
(OSA
37)
was
ligated
into
the
partially
digested
plasmid.
In
addition
to
the
uniqueApaI
site,
pDRO
contains
unique
SpeI
and
Bcll
sites
located
5'
and
3',
respectively,
to
the
sequences
coding
for
the
two
Zn2+
fingers.
Plasmid
pDR1
was
generated
by
deleting
a
423-bp
ApaI-
BclI
fragment
from
pDRO.
After
digestion
with
ApaI
and
BclI,
pDR0
was
religated
with an
oligonucleotide
with
the
sequence
5'-GAT
CAT
CGG
GCC-3'
(AR
5252),
which
was
used
as
a
cuttable
ApaI-BclI
adaptor.
Similarly,
plasmid
pDR2
was
constructed
by
removing
a
499-bp
SpeI-ApaI
fragment
from
pDRO.
The
SpeI-ApaI
sites
were
ligated
to
a
cuttable
oligonucleotide
adaptor
with
the
sequence
5'-CTA
GTA
CGG
GCC-3'
(AR
5253).
The
sequence
for
the
first
Zn2+
finger,
present
in
pDR1,
was
mutagenized
to
that
of
the
sequence
for
the
second
Zn2+
finger
in
the
HIV-1
NC
protein
by
using
a
mutagenic
oligonucleotide
with
the
sequence
5'-GGA
ACC
AAA
GAA
AGA
CTG
TTA
AGT
GIT
GGA
AAT
GTG
GAA
AGG
AAG
GAC ACC
AAA
TGA
AAG
ATT
GTA
GGG
CCC
GAT
GAT
CAG
ATA
CT-3'
(NAZ
21).
The
resulting
plas-
mid
is
designated
pDR4.
The
sequence
coding
for
the
second
Zn2+
finger
of
the
HIV-1
NC
protein,
present
in
pDR2,
was
mutated
to
that
of
the
first
by
using
a
mutagenic
oligonucle-
otide
with
the
sequence
5'-GGG
CCC
CTA
GGAAAA
AGG
GCT
GTT
TCA
ATT
GTG
GCA
AAG
AAG
GGC
ACA
TAG
CCA
AAA
ATT
GCA
CTG
AGA
GAC
AGG
CTA
AT-3'
(NAZ
20).
The
resulting
plasmid
is
designated
pDR6.
Plasmid
pDR7
contains
the
sequence
for
the
first
Zn2+
finger
in
both
the
first-
and
second-finger
positions
of
the
HIV-1
NC
protein.
This
plasmid
was
reconstructed
by
opening
pDR6
with
SpeI
and
ApaI
and
ligating
in
the
499-bp
SpeI-ApaI
fragment
from
pDR1.
Subclone
pDR8
contains
sequences
for
the
second
Zn2+
finger
in
both
the
first-
and
second-finger
positions.
pDR8
was
generated
by
cutting
pDR2
with
SpeI
and
ApaI
and
ligating
in
the
499-bp
SpeI-
ApaI
fragment
containing
the
sequence
for
the
second
Zn2+
finger
from
pDR4.
Plasmid
pDR9
is
a
subclone
in
which
the
positions
of
the
sequences
coding
for
first
and
second
Zn2+
fingers
are
reversed
with
respect
to
wild-type
virus.
pDR9
was
created
by
opening
pDR6
with
SpeI
and
ApaI
and
ligating
in
a
499-bp
SpeI-ApaI
fragment
from
pDR4.
Full-length
proviral
clones
were
reconstructed
by
ligating
the
4,278-bp
SpeI-SalI
fragments
from
subclones
pDR7,
pDR8,
and
pDR9
with
the
10.7-kbp
SpeI-SalI
fragment
from
pNL4-3
(Fig.
1).
Full-length
mutant
and
wild-type
clones
are
designated
as
follows.
pNC1/2
(pNL4-3)
is
the
wild-type
proviral
clone,
in
which
sequences
coding
for
the
first
and
second
Zn2+
fingers
are
in
the
first-
and
second-finger
positions,
respectively,
of
the
HIV-1
NC
protein.
pNC1/1
(from
pDR7)
and
pNC2/2
(from
pDR8)
are
the
mutants
with
the
duplication
of
the
first
or
second
Zn2+
finger
sequences,
J.
VIROL.

HIV-1
ZINC
FINGER
REARRANGEMENT
MUTANTS
4029
(pBluescript
KS+)
Spel/SaII
/
SaiI
Bc1I
Sal
BcilI
SalI
BclI
Sail
BclI
FIG.
1.
Schematic
diagram
of
HIV-1
NC
mutant
construction.
Plasmids
used
in
the
construction
of
the
NC
protein
mutants
are
indicated
by
the
type
(either
the
sequence
for
the
first
Zn2+-finger
[*1:
CFNCGKEGHIAKNC]
or
that
for
the
second
Zn2+
finger
[*2-
CWKCGKEGHQMKDC])
and
location
of
the
sequence
for
the
Zn2+-fingers.
DNA
fragments
and
oligonucleotides
used
for
constructions
are
indicated
in
parentheses.
Oligonucleotides
used
for
oligonucleotide-directed
mutagenesis
are
indicated
in
brackets.
Plasmids
and
restriction
sites
are
not
drawn
to
scale.
respectively,
in
the
first-
and
second-finger
positions
of
the
HIV-1
NC
protein.
pNC2/1
is
the
mutant
proviral
clone
with
the
positions
of
the
first
and
second
Zn2+
finger
sequences
reversed.
All
mutations
were
confirmed
by
direct
sequencing
with
Sequenase
(United
Stated
Biochemical
Corp.,
Cleve-
land,
Ohio).
Transfection.
Plasmids
were
transfected
into
80
to
90%
confluent
HeLa
or
HCLZ
cell
monolayers
by
the
calcium
phosphate
method
(11).
HeLa
cells
were
transfected
in
150-cm2
tissue
culture
flasks.
We
took
24-h
harvests
3
to
4
days
after
transfection.
Supernatants
were
clarified,
and
the
virus
was
analyzed.
HCLZ
cells
were
transfected
in
25-cm2
flasks
24
h
after
seeding
at
a
density
of
8
x
105
cells
per
flask.
HCLZ
cell
monolayers
were
developed
3
days
posttransfec-
tion
with
5-bromo-4-chloro-3-indolyl-3-D-galactoside
(X-Gal
reagent
[16])
to
locate
cells
containing
viral
constructs
pro-
ducing
HIV-1
Tat.
RT
assays.
Reverse
transcriptase
(RT)
assays
were
per-
formed
on
clarified
supernatants
from
transfections
and
infectivity
assays
as
described
previously
(10).
Assays
for
viral
proteins.
Virus
was
harvested
as
described
previously
(10).
Clarified
supernatants
were
analyzed
by
using
the
HIV
p24CA
antigen
capture
enzyme-linked
immu-
nosorbent
assay
kit
(DuPont/NEN
Research
Products,
Bos-
ton,
Mass.).
Samples
were
adjusted
for
equal
levels
of
p24CA
and
analyzed
as
follows.
Viral
proteins
were
fractionated
by
sodium
dodecyl
sulfate-polyacrylamide
gel
electrophoresis
(14)
and
transferred
to
Immobilon-P
filters
(Millipore
Corp.,
Bedford,
Mass.)
(30).
Specific
viral
proteins
were
detected
by
using
goat
antisera
to
p7NC,
p24CA,
and
gpl20Su.
The
immunotransfers
were
then
incubated
with
a
secondary
rabbit
anti-goat
horseradish
peroxidase-conjugated
antibody
(Bio-Rad).
Viral
proteins
were
visualized
by
using
the
en-
hanced
chemiluminescence
(ECL)
Immunodetection
System
(Amersham).
RNA
blot
analysis.
The
genomic
RNA
content
in
these
mutant
virions
was
analyzed
as
described
previously
(10).
Infectivity
assays.
Infectivity
assays
were
performed
with
H9
cells
as
described
previously
(10),
with
1
ml
of
inoculum
incorporating
2
,ug
of
Polybrene
per
ml.
Samples
from
the
VOL.
67,
1993

4030
GORELICK
ET
AL.
transfected
HeLa
cells
were
clarified
by
two
successive
centrifugation
steps.
Samples
from
infected
H9
cultures
were
taken
biweekly
and
tested
for
the
presence
of
virus
by
monitoring
supematant
RT
levels.
Limiting
dilution
of
virus
from
the
transfection
of
the
wild-type
clone
pNL4-3
was
performed
to
determine
the
concentration
of
infectious
par-
ticles
resulting
from
a
typical
transfection.
HCLZ
cells
were
seeded
into
25-cm2
flasks
e
1-nsity
of
8
x
105
cells
per
flask
and
infected
24
h
later
v,
.'
,utant
and
wild-type
virus
from
the
HeLa
cell
transfecti-
Then
2
ml
of
clarified
supernatant
from
transfected
I-
,La
cells
was
applied
to
the
HCLZ
cells
in
the
presence
of
2
,ug
of
Polybrene
per
ml.
HCLZ
cells
were
incubated
overnight
and
then
washed
twice
with
5
ml
of
Hanks'
balanced
salt
solution.
The
monolayers
were
then
incubated
for
an
addi-
tional
48
h
with
10
ml
of
medium
and
developed
as
described
previously
(16).
Electron
microscopy.
Electron
micrographs
were
per-
formed
by
K.
Nagashima,
Laboratory
of
Cellular
and
Mo-
lecular
Structure,
Program
Resources
Inc./DynCorp,
NCI-
FCRDC.
HCLZ
cells
transfected
with
mutant
and
wild-type
proviral
clones
were
fixed
in
2%
glutaraldehyde-2%
formal-
dehyde
and
developed
with
X-Gal
reagent,
embedded
di-
rectly
from
the
tissue
culture
flask,
and
processed
as
de-
scribed
previously
(8).
Blue
cells
were
selectively
sectioned
and
examined
in
Hitachi
H-7000
electron
microscope
oper-
ated
at
75
kV.
0
z
a)
0o
cn
a)
C)
S
a)H
S
a)
a)
cS
+
on
N
C)
ca
S
C)
._
CS
U,
._
U,
CS
._
U,
c0
RESULTS
Mutagenesis.
To
determine
whether
the
positions
of
the
two
Zn2+
fingers
of the
HIV-1
NC
protein
are
interchange-
able,
we
constructed
three
mutants
by
using
the
NY5/LAV
molecular
clone
of
HIV-1,
pNL4-3.
Subclones
containing
the
individual
Zn2+
fingers
were
constructed
and
mutated
by
oligonucleotide-directed
mutagenesis.
Full-length
proviral
clones
were
then
reconstructed
and
analyzed
(Fig.
1).
Table
1
summarizes
the
amino
acid
and
nucleotide
mutations
that
result
from
rearrangement
of
Zn2+
finger
sequences
in
the
mutant
clones.
As
indicated
in
Fig.
1
and
Table
1,
these
mutants
include
duplication
of
the
first
and
second
Zn2+
finger
sequence
as
well
as
reversal
of
these
sequences.
Mutagenesis
with
the
80-base
oligonucleotides
NAZ
20
and
NAZ
21
was
highly
efficient
(data
not
shown)
for
the
replacement
of
Zn-+
finger
coding
sequences
in
clones
pDR1
and
pDR2
(Fig.
1).
Protein
composition
of
Zn2+
finger
rearrangement
mutants.
Mutant
and
wild-type
plasmids
were
transfected
onto
HeLa
cell
monolayers.
At
4
days
posttransfection,
supernatants
were
examined
for
RT
activity
and
p24CA
content.
Table
2
shows
that
the
mutant
and
wild-type
clones
produce
similar
levels
of
RT
and
p24',
as
was
observed
previously
for
other
NC
mutants
(2,
10).
Immunoblot
analysis
performed
on
these
mutants
reveals
the
presence
of
properly
processed
p24CA,
p7NC,
and
gpl20SU
when
compared
with
wild-type
virus
and
the
MN
strain
of
HIV-1
(Fig.
2).
These
results
(Table
2;
Fig.
2)
indicate
thatpol
gene
products
protease
and
RT
are
being
expressed
and
are
functioning
properly.
Sam-
ples
from
the
HeLa
cell
transfections
were
adjusted
for
equal
levels
of
p24CA.
The
intensities
of
the
bands
corresponding
to
the
proteins
analyzed
are
very
similar
for
both
the
Gag
and
Env
proteins.
There
are
slight
differences
in
the
mobility
of
the
p7
c,
which
may
be
due
to
differences
in
the
binding
of
sodium
dodecyl
sulfate.
HCLZ
cell
transfections
and
analysis.
To
examine
the
level
of
Tat
protein
production
and
syncytium
formation,
we
Ct
u
a)
4)
()
.o
0
0
S
r.
0
0
ca
0
n
tzI
z;
PI
05
¢
¢
0
0
0
0O
0x
0)
0)I
I
I
E
F1
A
1,:
1GP
GPI
C)
E-'
0
0
E-'0
4E
01-
0I
0
0i
4
El
0'
Eq
0
i
00
OH
E¢
H
0
i
cH
H
H
0
E-
¢
1
0
¢
I
i0
4I
10
I~
cI
10p
¢
EI
0z
0z
H i
IzE
EI
¢I
¢1
I)
I
0
0
44
0
I
0
H
H
I
IO
Hs
.0a
)
.'2a
ru_
S-
-
°
1--
Sh
43
O.M"
'U3
m
m
zI
J.
VIROL.
c)
._
C)
S
.2
.2
S
0
'.
0
I
S
'0

HIV-1
ZINC
FINGER
REARRANGEMENT
MUTANTS
4031
TABLE
2.
Characteristics
of
HIV-1
Zn2+
finger
rearrangement
mutants
Virus
Amt
of
RT
Amt
of
p24CA
pNC1/1
654,200
67,691
pNC2/2
341,720
66,835
pNC2/1
616,360
84,818
pNILA-3
(wild
type)
1,035,590
63,695
None
(calf
thymus
DNA)C
0
0
a
cpm
of
[3H]TrP
incorporated
per
milliliter
of
culture
fluid.
b
Picograms
of
p24CA
per
milliliter
of
culture
fluid
measured
by
p24CA
antigen
capture.
c
Cells
transfected
with
carrier
DNA
alone.
A
background
of
1,081
cpm/0.1
ml
has
been
subtracted
from
the
RT
values
shown.
transfected
HCLZ
cells
with
mutant
and
wild-type
clones.
All
mutant
and
the
wild-type
clones
were
Tat
positive
as
indicated
by
the
presence
of
0-galactosidase
production
in
the
HCLZ
cells;
approximately
1
to
5%
of
the
cells
in
the
monolayer
produced
Tat.
Additionally,
there
was
substantial
syncytium
formation
in
the
Tat-positive
HCLZ
cells
with
all
clones
tested.
This
indicates
that
functional
viral
envelope
proteins
are
expressed.
Electron-micrographic
analysis.
HCLZ
cells
were
ex-
tremely
useful
for
locating
cells
that
produce
virus.
Previous
attempts
to
obtain
electron
micrographs
of
mutant
HIV-1
particles
from
thin
sections
of
transfected
HeLa
cells
proved
exceedingly
difficult
because
of
the
small
numbers
of
cells
that
were
actually
producing
virus
(1
to
5%).
Virus-produc-
ing
HCLZ
cells
are
easily
located
as
visible,
P-galactosidase-
positive,
blue
syncytia
and
cells.
Thin
sections
of
transfected,
,B-galactosidase-positive
HCLZ
cells
were
examined
at
a
magnification
of
x
90,000
by
electron
microscopy.
Zn2+
finger
mutant
clones
produced
virion
particles
that
have
morphologies
identical
to
particles
obtained
from
the
wild-type
clone.
Representative
examples
showing
intra-
and
extracellular
mature
and
immature
parti-
cles
are
presented
in
Fig.
3.
Virus
particles
were
found
mainly
in
and
around
large
syncytia.
Particles
within
these
cells
were
located
in
intracellular
vacuoles.
HCLZ
cells
that
z
z
z
CL
CL
gp120Su
-_6
`
p24CC.
p7NC
t
-
-
d
kDa
116
97
31
14
6.5
FIG.
2.
Protein
immunoblotting
analysis
of
mutant
and
wild-type
virus
particles.
Samples
were
isolated
as
described
previously
(10).
All
samples
were
adjusted
for
an
equal
2.1
,ug
of
p24"A,
fractionated,
and
treated
as
described
in
Materials
and
Methods.
Molecular
masses
of
marker
proteins
are
indicated
on
the
right.
The
positions
of
p7NC,
p24CA,
and
gp120Su
are
indicated
on
the
left.
(-),
samples
from
HeLa
cells
transfected
with
calf
thymus
carrier
DNA.
were
transfected
with
calf
thymus
carrier
DNA
showed
no
blue
cells
on
development
with
X-Gal
reagent
(16),
and
electron
micrographs
were
negative
for
virus
particles,
as
expected
(Fig.
3).
HIV-1
NC
mutant
viruses
described
in
a
previous
study
(10)
were
examined
by
this
method,
and
similar
morphologies
were
observed
(data
not
shown).
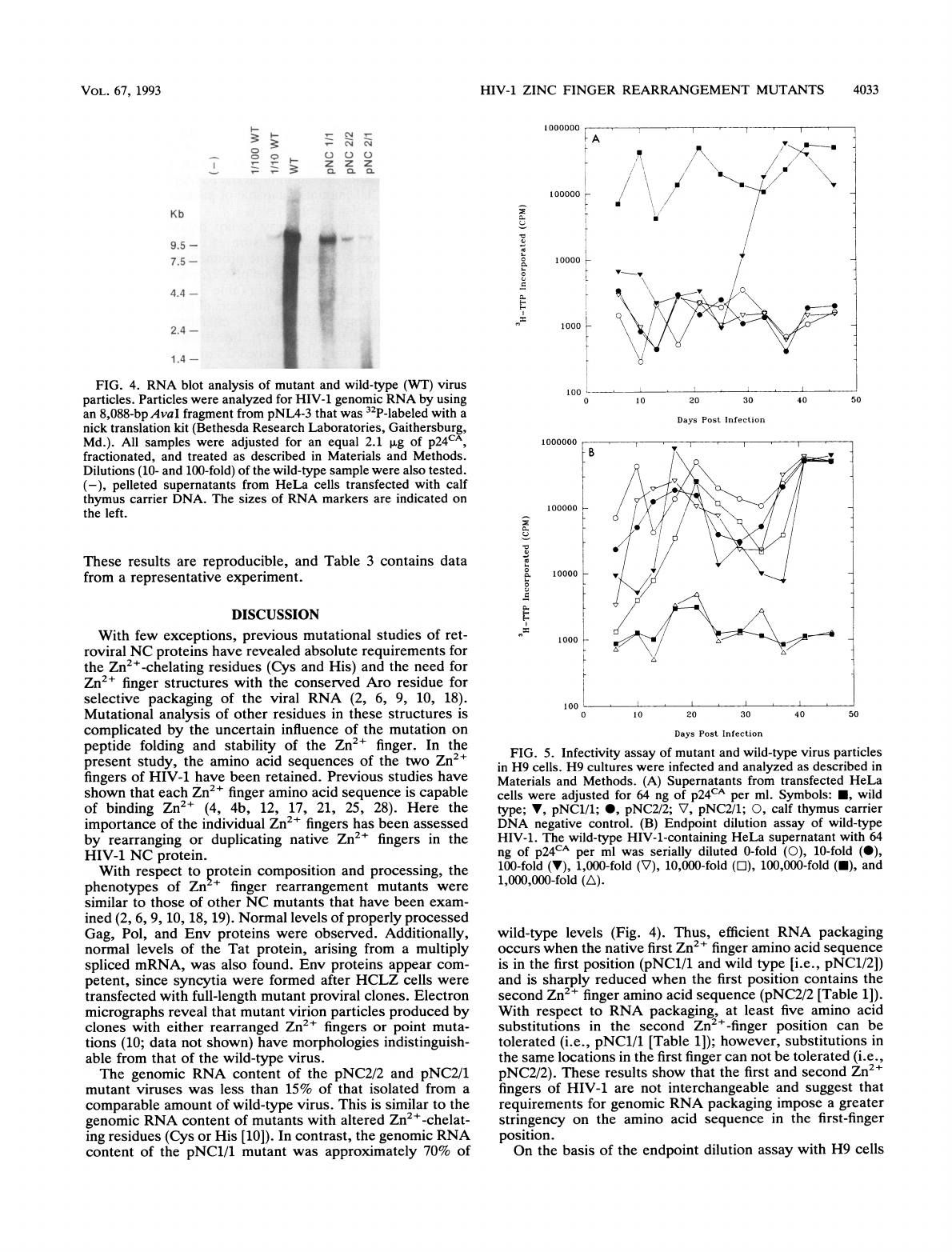
Genomic
RNA
content.
Viruses
from
the
mutant
and
wild-type
clones
were
adjusted
for
equal
amounts
of
p24CA,
and
the
RNA
was
extracted.
Figure
4
shows
results
of
a
Northern
(RNA)
analysis
performed
on
the
genomic
RNA
from
these
samples,
with
dilutions
of
the
wild-type
sample
to
calibrate
the
analysis.
The
pNC2/1
and
pNC2/2
mutants
package
approximately
10
and
15%
of
the
wild-type
levels
of
genomic
RNA
respectively,
which
is
similar
to
levels
found
in
other
HIV-1
NC
mutants
(10).
The
pNC1/1
mutant
con-
tains
approximately
70%
of
the
amount
of
genomic
RNA
found
in
wild-type
virus;
this
is
significantly
greater
than
had
been
observed
previously
with
other
Zn2+
finger
mutants
of
HIV-1
(2,
10).
Infectivity
analysis.
Wild-type
and
mutant
viruses
from
the
HeLa
cell
transfections
were
tested
for
their
ability
to
infect
H9
or
HCLZ
cells.
Results
from
infection
of
H9
cells
are
summarized
in
Fig.
5A.
All
inocula
were
adjusted
to
64
ng
of
p24CA.
After
incubation
of
wild-type
virus
with
H9
cells,
there
was
a
very
rapid
rise
in
RT
activity.
The
pNC1/1
mutant
appears
to
replicate
rapidly
after
a
lag
period
of
3
to
4
weeks.
This
phenotype
was
observed
with
two
replicate
clones.
The
pNC2/2
and
pNC2/1
mutants
are
noninfectious
(Fig.
5A).
An
endpoint
dilution
assay
was
performed
to
quantitate
the
relative
infectivity
of
the
wild-type
virus
(Fig.
SB).
Wild-type
virus
with
a
starting
p24CA
concentration
of
64
ng/ml
remained
infectious
after
10,000-fold
but
not
100,000-
fold
dilution.
Therefore,
the
titer
is
between
104
and
105
tissue
culture
infectious
doses
per
ml.
The
data
also
show
that
wild-type
virus
in
the
limiting
1:10,000
dilution
requires
approximately
2
weeks
to
spread
through
the
H9
culture
(Fig.
SB).
At
an
initial
inoculum
of
64
ng
of
p24CA
per
ml,
the
pNC1/1
mutant
requires
a
longer
incubation
period
before
spreading
rapidly
through
the
culture
(Fig.
SA).
The
sharp
rise
in
RT
activity
occurred
at
about
25
days
postinoculation
(pNC1/1;
Fig.
SA).
The
long
lag
period
and
sharp
rise
in
RT
activity
suggest
that
the
original
pNC1/1
mutant
may
have
reverted
to
a
wild-type
phenotype.
In
a
similar
experiment,
the
infectivity
profile
of
pNC1/1
virus
that
had
spread
throughout
an
H9
culture
(pNC1/1-H9)
was
examined.
The
same
amount
of
wild-type
virus
was
used
as
a
comparison.
The
clarified
supernatants
were
used
to
infect
naive
H9
cells.
As
seen
in
Fig.
6,
pNC1/1-H9
replicates
with
the
same
kinetics
as
that
of
the
wild-type
virus.
This
is
in
contrast
to
the
3.5-week
lag
period
seen
with
pNC1/1
virus
derived
from
HeLa
cell
transfections
(Fig.
SA).
These
results
support
the
hypothesis
that
the
pNC1/1
virus
replicates
at
an
undetectable
level
in
H9
cells
but
can
revert
to
a
competent,
highly
replicative
species
(e.g.,
pNC1/
1-H9).
It
should
be
noted
that
the
observations
with
respect
to
the
kinetics
of
infectivity
of
the
pNC1/1
mutant
are
reproducible.
HCLZ
cells
were
infected
with
mutant
and
wild-type
virus
from
HeLa
cell
transfections.
Table
3
shows
that
wild-type
virus
readily
infects
HCLZ
cells,
as
determined
by
the
number
of
3-galactosidase-producing
foci.
pNC2/2
and
pNC2/1
appear
to
be
able
to
infect
these
cells
but
only
at
a
very
low
frequency.
The
pNC1/1
mutant
is
able
to
infect
HCLZ
cells
at
a
level
of
10%
of
that
of
wild-type
virus.
VOL.
67,
1993

4032
GORELICK
ET
AL.
CZ
C:
*
i
-A-L~~~~~~~~~~~~~~~~~~~~.
CL8_
LC
Z0
.~~~~~~~~~~~~~~~~~~~~~~.
.,=
CM)
04
0~~~~
a)
0
i
~
.
CZ
C.
I
iI
ti-sss
Ee
z
o~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~Z
0.~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~C
oQ
:a
l
K
{
;fw44+SS
F
>
=~~~~~~~~~~~~I.
}L;
0
<
CM
u,~~~~~~0
CL
t
il-
--
e-
i88t . 5 3 ;t ;t
0
o
.
W
>
1
o
?f
,
*
Xjp
P
~Z
X
*
e
i
SW
i
E
M
<
b
Q
C,
~~~~~~u
0
J
^
_
.
f
-
*
q
_
&
*
'
t
Q
J.
VIROL.
6wh"
A".''
*-".5,
HIV-1
ZINC
FINGER
REARRANGEMENT
MUTANTS
4033
1000000
C)
C)
C)
z
zz
Q.L
0.
0.
Kb
9.5
-
7.5
-
4.4
-
2.4
-
1.4-
_
"I
1
00000
10000
1000
FIG.
4.
RNA
blot
analysis
of
mutant
and
wild-type
(WT)
virus
particles.
Particles
were
analyzed
for
HIV-1
genomic
RNA
by
using
an
8,088-bpAvaI
fragment
from
pNL4-3
that
was
32P-labeled
with
a
nick
translation
kit
(Bethesda
Research
Laboratories,
Gaithersburg,
Md.).
All
samples
were
adjusted
for
an
equal
2.1
pLg
of
p24CA,
fractionated,
and
treated
as
described
in
Materials
and
Methods.
Dilutions
(10-
and
100-fold)
of
the
wild-type
sample
were
also
tested.
(-),
pelleted
supernatants
from
HeLa
cells
transfected
with
calf
thymus
carrier
DNA.
The
sizes
of
RNA
markers
are
indicated
on
the
left.
These
results
are
reproducible,
and
Table
3
contains
data
from
a
representative
experiment.
DISCUSSION
With
few
exceptions,
previous
mutational
studies
of
ret-
roviral
NC
proteins
have
revealed
absolute
requirements
for
the
Zn2+-chelating
residues
(Cys
and
His)
and
the
need
for
Zn2+
finger
structures
with
the
conserved
Aro
residue
for
selective
packaging
of
the
viral
RNA
(2,
6,
9,
10,
18).
Mutational
analysis
of
other
residues
in
these
structures
is
complicated
by
the
uncertain
influence
of
the
mutation
on
peptide
folding
and
stability
of
the
Zn2+
finger.
In
the
present
study,
the
amino
acid
sequences
of
the
two
Zn2+
fingers
of
HIV-1
have
been
retained.
Previous
studies
have
shown
that
each
Zn2+
finger
amino
acid
sequence
is
capable
of
binding
Zn2+
(4,
4b,
12,
17,
21,
25,
28).
Here
the
importance
of
the
individual
Zn2+
fingers
has
been
assessed
by
rearranging
or
duplicating
native
Zn2+
fingers
in
the
HIV-1
NC
protein.
With
respect
to
protein
composition
and
processing,
the
phenotypes
of
Zn
+
finger
rearrangement
mutants
were
similar
to
those
of
other
NC
mutants
that
have
been
exam-
ined
(2,
6, 9,
10,
18,
19).
Normal
levels
of
properly
processed
Gag,
Pol,
and
Env
proteins
were
observed.
Additionally,
normal
levels
of
the
Tat
protein,
arising
from
a
multiply
spliced
mRNA,
was
also
found.
Env
proteins
appear
com-
petent,
since
syncytia
were
formed
after
HCLZ
cells
were
transfected
with
full-length
mutant
proviral
clones.
Electron
micrographs
reveal
that
mutant
virion
particles
produced
by
clones
with
either
rearranged
Zn2+
fingers
or
point
muta-
tions
(10;
data
not
shown)
have
morphologies
indistinguish-
able
from
that
of
the
wild-type
virus.
The
genomic
RNA
content
of
the
pNC2/2
and
pNC2/1
mutant
viruses
was
less
than
15%
of
that
isolated
from
a
comparable
amount
of
wild-type
virus.
This
is
similar
to
the
genomic
RNA
content
of
mutants
with
altered
Zn2+-chelat-
ing
residues
(Cys
or
His
[10]).
In
contrast,
the
genomic
RNA
content
of
the
pNC1/1
mutant
was
approximately
70%
of
10o
1000000
5-
Iz
S.
0
In
100000
10000
1000
100
0
0
1
0
20
30
Days
Post
Infection
10
20
30
Days
Post
Infection
40
50
40
50
FIG.
5.
Infectivity
assay
of
mutant
and
wild-type
virus
particles
in
H9
cells.
H9
cultures
were
infected
and
analyzed
as
described
in
Materials
and
Methods.
(A)
Supernatants
from
transfected
HeLa
cells
were
adjusted
for
64
ng
of
p24CA
per
ml.
Symbols:
*,
wild
type;
V,
pNC1/1;
*,
pNC2/2;
V,
pNC2/1;
0,
calf
thymus
carrier
DNA
negative
control.
(B)
Endpoint
dilution
assay
of
wild-type
HIV-1.
The
wild-type
HIV-1-containing
HeLa
supernatant
with
64
ng
of
p24CA
per
ml
was
serially
diluted
0-fold
(0),
10-fold
(0),
100-fold
(V),
1,000-fold
(V),
10,000-fold
([),
100,000-fold
(-),
and
1,000,000-fold
(A).
wild-type
levels
(Fig.
4).
Thus,
efficient
RNA
packaging
occurs
when
the
native
first
Zn2+
finger
amino
acid
sequence
is
in
the
first
position
(pNC1/1
and
wild
type
[i.e.,
pNC1/2])
and
is
sharply
reduced
when
the
first
position
contains
the
second
Zn2+
finger
amino
acid
sequence
(pNC2/2
[Table
1]).
With
respect
to
RNA
packaging,
at
least
five
amino
acid
substitutions
in
the
second
Zn2+-finger
position
can
be
tolerated
(i.e.,
pNC1/1
[Table
1]);
however,
substitutions
in
the
same
locations
in
the
first
finger
can
not
be
tolerated
(i.e.,
pNC2/2).
These
results
show
that
the
first
and
second
Zn2+
fingers
of
HIV-1
are
not
interchangeable
and
suggest
that
requirements
for
genomic
RNA
packaging
impose
a
greater
stringency
on
the
amino
acid
sequence
in
the
first-finger
position.
On
the
basis
of
the
endpoint
dilution
assay
with
H9
cells
A~
I
~~/
.0
B
VOL.
67,
1993
i.
CL
"I
co
L.
a
6.0.
0
u
.E
a.
E--
E-
0

4034
GORELICK
ET
AL.
1000000
100000
R
0
I"I
0-
E-
10000
1000
100
0
10
20
30
40
Days
Post
Infection
FIG.
6.
Infectivity
assay
of
pNC1/1,
pNC1/1-H9,
and
wild-type
virus
particles
in
H9
cells.
H9
cultures
were
infected
and
analyzed
as
described
in
Materials
and
Methods.
Supernatants
containing
18
ng
of
p24CA
per
ml
for
pNC1/1-H9
(-)
and
wild-type
HIV-1
(A)
were
used
to
infect
H9
cells.
The
pNC1/1
curve
(with
a
starting
inoculum
of
64
ng/ml)
from
Fig.
5A
(-)
is
shown
for
comparison.
(Fig.
SB),
the
pNC2/2
and
pNC2/1
viruses
have
specific
infectivities
that
are
at
least
104-fold
lower
than
that
of
wild-type
virus.
This
phenotype
is
similar
to
the
phenotype
of
other
Zn2+
finger
mutants
with
altered
Zn2+-chelating
residues
(10)
and
is
in
keeping
with
their
low
content
of
genomic
RNA.
Although
the
pNCl/1
mutant
is
capable
of
infecting
H9
cells,
the
kinetics
of
infection
(Fig.
5A)
is
not
indicative
of
a
low
titer
of
virus
as
is
seen
for
a
10,000-fold
dilution
of
wild-type
virus
(Fig.
SB).
For
the
pNC1/1
mutant,
there
is
a
lag
period
of
3
to
4
weeks
followed
by
a
rapid
rise
in
RT
activity.
Virus
taken
from
the
culture
at
the
peak
of
RT
activity
(pNC1/1-H9
virus)
is
able
to
infect
naive
H9
cells
as
readily
as
wild-type
virus
does
(Fig.
6).
This
suggests
that
the
pNC1/1-H9
virus
is
different
from
the
original
pNC1/1
mutant
and
that
the
pNC1/1-H9
virus
may
have
reverted
to
a
wild-type
phenotype
through
mutations,
either
in
the
second
Zn2+
finger
sequence
or
elsewhere.
Similar
results
to
these
were
observed
by
Rein
et
al.
(24)
for
a
replication-
defective
Moloney
murine
leukemia
virus.
These
results
also
TABLE
3.
Infectivity
assay
of
HIV-1
Zn2+
finger
rearrangement
mutants
with
HCLZ
cells
Amt
of
p24A
Avg
Corrected
Virus
in
inoculum
no.
of
no.
of
(ng/ml)a
focib
focic
pNC1/1
107.4
3.4
1.5
pNC2/2
54.8
0.9
0.5
pNC2/1
86.0
0.8
0.3
pNL4-3
(wild
type)
98.0
27.1
14.9
None
(calf
thymus
DNA)
0.0
0.4
0.0
a
p24CA
content
of
clarified
supernatants
from
HeLa
cell
transfections
was
measured
by
p24CA
antigen
capture.
b
Average-focus
values
were
determined
by
counting
the
number
of
f-ga-
lactose-positive
cells
per
field
with
a
1Ox
eyepiece
and
objective.
A
total
of
30
fields
were
counted,
and
the
values
were
averaged.
c
The
corrected-foci
values
were
determined
by
first
subtracting
the
back-
ground
average-focus
value
for
calf
thymus
DNA
from
the
average-focus
value
and
then
adjusting
for
a
constant
level
of
54.8
ng
of
p24CA
per
ml.
suggest
that
the
pNC1/1
mutant
is
replication
deficient
and
probably
sustains
a
nondetectable
level
of
infection
in
H9
cells
until
this
reversion
occurs.
However,
the
replication
deficiency
does
not
appear
to
be
at
the
RNA-packaging
step,
since
the
pNC1/1
mutant
is
capable
of
packaging
viral
RNA
to
at
least
70%
of
wild-type
levels.
The
HCLZ
cell
focus
assay
is
not
dependent
on
viral
replication
and
provides
a
measure
of
early
events
in
the
infectious
process,
leading
to
the
production
of
the
regula-
tory
Tat
protein.
The
pNC2/1
and
pNC2/2
mutants
gave
very
low
levels
of
foci
in
this
assay
(Table
3),
which
were
not
significantly
different
from
the
numbers
of
foci
obtained
with
other
NC
mutants
with
altered
Zn2+-chelating
residues
(Cys
to
Ser
mutants
[data
not
shown]).
Infections
of
HCLZ
cells
with
the
pNC1/1
mutant
produced
a
larger
number
of
foci
than
infections
with
the
pNC2/2
and
pNC2/1
mutants,
but
the
number
of
foci
was
only
10%
of
that
obtained
by
infections
with
wild-type
virus.
Therefore,
even
though
levels
of
RNA
packaged
in
the
pNC1/1
mutant
were
compa-
rable
to
wild-type
levels,
the
ability
of
this
mutant
to
score
positive
in
this
assay
was
only
10%
of
that
of
the
wild
type
(Table
3).
These
results
show
that
the
pNC1/1
mutant
is
inefficient
in
its
ability
to
carry
out
early
steps
in
the
infectious
process
that
lead
to
the
production
of
viral
pro-
teins.
This
phenotype
strongly
suggests
that,
in
addition
to
their
role
in
viral
RNA
packaging,
the
Zn2+
fingers
of
Gag
precursors
and
NC
proteins
are
required
for
critical
pro-
cesses
in
the
early
stages
of
infection.
We
previously
sug-
gested
such
an
additional
role
for
the
NC
protein
based
on
the
phenotype
of
NC
mutants
derived
from
Moloney
murine
leukemia
virus
(9).
The
exact
role
of
the
NC
protein
in
the
early
stages
of
infection
cannot
be
deduced
from
the
data
provided
here;
however,
some
possibilities
can
be
excluded.
Transfections
of
HCLZ
cells
with
proviral
DNA
containing
mutations
in
the
NC
protein
(pNC1/1,
pNC2/1,
pNC2/2,
or
Zn2+-chelating
residues)
produce
as
many
foci
as
are
ob-
tained
by
transfections
with
wild-type
proviral
DNA
(data
not
shown).
Therefore,
the
NC
mutants
do
not
appear
deficient
in
events
that
follow
DNA
production
and
lead
to
protein
synthesis.
However,
viral
entry
and
reverse
tran-
scription
of
viral
RNA
are
early
steps
in
the
infectious
process
that
could
involve
the
NC
protein.
Our
data
do
not
distinguish
between
these
possibilities,
but
others
have
suggested
roles
for
the
NC
protein
in
primer
tRNA
binding
and
reverse
transcription
of
viral
RNA
(7,
23).
Our
results
are
consistent
with
this
suggestion.
Physical
chemistry
studies
have
shown
that
the
NC
pro-
tein
binds
to
single-stranded
nucleotides
(DNA
or
RNA)
without
great
specificity
for
a
given
nucleotide
sequence
(13,
28).
It
probably
uses
this
general
polynucleotide-binding
property
in
the
core
of
the
mature
virus,
where
2,000
to
3,000
NC
proteins
may
bind
to
the
two
single
strands
of
genomic
RNA
(4,
13).
However,
genomic
RNA
packaging
requires
that
there
be
a
specificity
for
some
feature
of
the
viral
RNA
that
is
superimposed
on
the
general
nucleotide-binding
prop-
erty
of the
NC
domain
in
the
Gag
precursor.
The
data
provided
here
for
HIV-1
indicate
that
some
or
all
of
the
specific
amino
acid
residues
at
the
Aro,
Xb,
Xg,
Xh,
and
Car
positions
(Table
1)
in
the
first
finger
are
required
for
specific
viral
RNA
packaging
(i.e.,
pNC2/2
versus
pNC1/2
[wild
type]
and
pNC1/1).
The
three-dimensional
structure
of the
HIV-1
NC
protein
and
its
Zn2+
fingers
has
been
determined
by
high-resolution
nuclear
magnetic
resonance
spectroscopy
(21,
25,
28,
29).
Nuclear
magnetic
resonance
studies
show
that
for
each
Zn2+
finger,
residues
at
positions
Aro,
Xb,
Xg,
and
Xh
form
part
of
a
hydrophobic
groove
that
is
believed
to
J.
VIROL.

HIV-1
ZINC
FINGER
REARRANGEMENT
MUTANTS
4035
interact
directly
with
nucleotide
bases
in
the
bound
state
(27,
28).
This
indicates
that
these
specific
residues
in
the
first
finger
may
play
a
prominent
role
in
the
recognition
and
packaging
of
viral
RNA
by
contributing
additional
stabilizing
interactions
with
specific
nucleotides.
However,
we
do
not
suggest
that
these
stabilizing
interactions
are
sufficient
to
direct
specific
recognition
of
genomic
RNA,
and
it
seems
probable
that
other
properties
of
the
Gag
precursor
and
RNA
play
prominent
roles
in
this
selection
and
packaging
process.
The
data
provided
here
extend
our
understanding
of
the
critical
roles
played
by
NC
Zn2+
fingers
in
the
replication
cycle
of
retroviruses.
In
the
Gag
precursor,
these
structures
are
essential
for
viral
RNA
packaging
during
assembly.
In
the
mature
virus,
the
same
structures
are
required
for
the
proper
functioning
of
the
NC
protein
in
early
stages
of
the
infectious
process.
It
seems
reasonable
to
suggest
that,
for
most
retroviruses,
the
dual
roles
performed
by
these
struc-
tures
have
led
to
the
evolutionary
development
of
two
distinct
Zn2+
fingers
per
NC
protein.
In
any
event,
the
finding
that
the
highly
conserved
Zn2+
fingers
in
the
NC
protein
operate
in
the
early
stages
of
the
infectious
process
has
important
implications
for
the
development
of
antiviral
drugs
to
specifically
attack
these
structure
in
the
mature
virus.
ACKNOWLEDGMENTS
We
thank
Kunio
Nagashima
for
assistance
in
providing
electron
micrographs
of
these
mutant
viruses,
Donald
G.
Johnson
for
assis-
tance
with
Western
analyses,
Lisa
D.
Stevenson
for
assistance
with
p24CA
quantitation,
and
Patricia
C.
Grove
for
assistance
in
prepa-
ration
of
the
manuscript.
This
research
was
sponsored
in
part
by
the
National
Cancer
Institute
under
contract
N01-CO-74102
with
Program
Resources,
Inc.,
and
N01-CO-74101
with
Advanced
BioScience
Laboratories,
Inc.
ADDENDUM
After
this
paper
was
submitted,
a
report
appeared
imply-
ing
that
cysteine
arrays
of
avian
retroviruses
function
in
stabilization
rather
than
packaging
of
viral
RNA
(2a).
This
alternative
interpretation
is
also
consistent
with
our
data
on
HIV-1.
Additionally,
since
submission
of
this
paper,
a
recent
report
by
Bowles
et
al.
(4a)
appeared
describing
the
effects
of
rearranging
the
Zn2+
fingers
of
Rous
sarcoma
virus.
The
results
from
our
study
are
generally
in
agreement
with
those
of
Bowles
et
al.
REFERENCES
1.
Adachi,
A.,
H.
E.
Gendelman,
S.
Koenig,
T.
Folks,
R.
Willey,
A.
Rabson,
and
M.
A.
Martin.
1986.
Production
of
acquired
immu-
nodeficiency
syndrome-associated
retrovirus
in
human
and
non-
human
cells
transfected
with
an
infectious
molecular
clone.
J.
Virol.
59:284-291.
2.
Aldovini,
A.,
and
R.
A.
Young.
1990.
Mutations
of
RNA
and
protein
sequences
involved
in
human
immunodeficiency
virus
type
1
packaging
result
in
production
of
noninfectious
virus.
J.
Virol.
64:1920-1926.
2a.Aronoff,
R.,
A.
M.
Hajjar,
and
M.
L.
Linial.
1993.
Avian
retroviral
RNA
encapsidation:
reexamination
of
functional
5'
RNA
sequences
and
the
role
of
nucleocapsid
Cys-His
motifs.
J.
Virol.
67:178-188.
3.
Berg,
J.
1986.
Potential
metal-binding
domains
in
nucleic
acid
binding
proteins.
Science
232:485-487.
4.
Bess,
J.
W.,
Jr.,
P.
J.
Powell,
H.
J.
Issaq,
L.
J.
Schumack,
M.
K.
Grimes,
L.
E.
Henderson,
and
L.
0.
Arthur.
1992.
Tightly
bound
zinc
in
human
immunodeficiency
virus
type
1,
human
T-cell
leukemia
virus
type
I,
and
other
retroviruses.
J.
Virol.
66:840-
847.
4a.Bowles,
N.
E.,
P.
Damay,
and
P.-F.
Spahr.
1993.
Effects
of
rearrangements
and
duplications
of
the
Cys-His
motifs of
Rous
sarcoma
virus
nucleocapsid
protein.
J.
Virol.
67:623-631.
4b.Chance,
M.
R.,
I.
Sagi,
M.
D.
Wirt,
S.
M.
Frisbie,
E.
Scheuring,
E.
Chen,
J.
W.
Bess,
Jr.,
L. E.
Henderson,
L.
0.
Arthur,
T.
L.
South,
G.
Perez-Alvarado,
and
M.
F.
Summers.
1992.
Extended
x-ray
absorption
fine
structure
studies
of
a
retrovirus:
equine
infectious
anemia
virus
cysteine
arrays
are
coordinated
to
zinc.
Proc.
Natl.
Acad.
Sci.
USA
89:10041-10045.
5.
Covey,
S.
N.
1986.
Amino
acid
sequence
homology
in
gag
region
of
reverse
transcribing
elements
and
the
coat
protein
gene
of
cauliflower
mosaic
virus.
Nucleic
Acids
Res.
14:623-633.
6.
Dupraz,
P.,
S.
Oertle,
C.
Mirc,
P.
Damay,
and
P.-F.
Spahr.
1990.
Point
mutations
in
the
proximal
Cys-His
box
of
Rous
sarcoma
virus
nucleocapsid
protein.
J.
Virol.
64:4978-4987.
7.
Khan,
R.,
and
D.
P.
Giedroc.
1992.
Recombinant
human
immu-
nodeficiency
virus
type
I
nucleocapsid
(NCP7)
protein
unwinds
tRNA.
J.
Biol.
Chem.
267:6689-6695.
8.
Gonda,
M.
A.,
S.
A.
Aaronson,
N.
Ellmore,
V.
H.
Zeve,
and
K.
Nagashima.
1976.
Ultrastructural
studies
of
surface
features
of
human
normal
and
tumor
cells
in
tissue
culture
by
scanning
and
transmission
electron
microscopy.
J.
Natl.
Cancer
Inst.
56:245-
263.
9.
Gorelick,
R.
J.,
L.
E.
Henderson,
J.
P.
Hanser,
and
A.
Rein.
1988.
Point
mutants
of
Moloney
murine
leukemia
virus
that
fail
to
package
viral
RNA:
evidence
for
specific
RNA
recognition
by
a
"zinc
finger-like"
protein
sequence.
Proc.
Natl.
Acad.
Sci.
USA
85:8420-8424.
10.
Gorelick,
R.
J.,
S.
M.
Nigida,
Jr.,
J.
W.
Bess,
Jr.,
L.
0.
Arthur,
L.
E.
Henderson,
and
A.
Rein.
1990.
Noninfectious
human
immunodeficiency
virus
type
1
mutants
deficient
in
genomic
RNA.
J.
Virol.
46:3207-3211.
11.
Graham,
F.
L.,
and
A.
J.
van
der
Eb.
1973.
A
new
technique
for
the
assay
of
infectivity
of
human
adenovirus
5
DNA.
Virology
52:456-467.
12.
Green,
L.
M., and
J.
M.
Berg.
1989.
A
retroviral
Cys-Xaa2-Cys-
Xaa4-His-Xaa4-Cys
peptide
binds
metal
ions:
spectroscopic
studies
and
a
proposed
three-dimensional
structure.
Proc.
Natl.
Acad.
Sci.
USA
86:4047-4051.
13.
Karpel,
R.
L.,
L. E.
Henderson,
and
S.
Oroszlan.
1987.
Interac-
tions
of
retroviral
structural
proteins
with
single-stranded
nu-
cleic
acids.
J.
Biol.
Chem.
162:4961-4967.
14.
Laemmli,
U.
K.
1970.
Cleavage
of
structural
proteins
during
the
assembly
of
the
head
of
bacteriophage
T4.
Nature
(London)
227:680-685.
15.
Leis,
J.,
D.
Baltimore,
J.
M.
Bishop,
J.
Coffin,
E.
Fleissner,
S.
P.
Goff,
S.
Oroszlan,
H.
Robinson,
A.
M.
Skalka,
H.
M.
Temin,
and
V.
Vogt.
1988.
Standardized
and
simplified
nomenclature
for
proteins
common
to
all
retroviruses.
J.
Virol.
62:1808-1809.
16.
MacGregor,
G.
R.,
G.
P.
Nolan,
S.
Fiering,
M.
Roederer,
and
L.
A.
Herzenberg.
1991.
Use
of
E.
coli
lacZ
(p-galactosidase)
as
a
reporter
gene,
p.
217-235.
In
E.
J.
Murray
and
J.
M.
Walker
(ed.),
Methods
in
molecular
biology,
vol.
7.
Gene
transfer
and
expression
protocols.
Humana
Press
Inc.
Clifton,
N.J.
17.
Mely,
Y.,
F.
Cornille,
M.-C.
Fournie-Zaluski,
J.-L.
Darlix,
B.
P.
Roques,
and
D.
Gerard.
1991.
Investigation
of
zinc-binding
affinities
of
Moloney
murine
leukemia
virus
nucleocapsid
pro-
tein
and
its
related
zinc
finger
and
modified
peptides.
Biopoly-
mers
31:899-906.
18.
Meric,
C.,
and
S.
P.
Goff.
1989.
Characterization
of
Moloney
murine
leukemia
virus
mutants
with
single-amino-acid
substitu-
tions
in
the
Cys-His
box
of
the
nucleocapsid
protein.
J.
Virol.
63:1558-1569.
19.
Meric,
C.,
E.
Gouilloud,
and
P.-F.
Spahr.
1988.
Mutations
in
Rous
sarcoma
virus
nucleocapsid
protein
p12
(NC):
deletions
of
Cys-His
boxes.
J.
Virol.
62:3328-3333.
20.
Meyers,
G.,
B.
Korber,
J.
A.
Berzofsky,
R.
F.
Smith,
and
G.
N.
Pavlakis.
1991.
Human
retroviruses
and
AIDS
1991.
A
compi-
lation
and
analysis
of
nucleic
acid
and
amino
acid
sequences,
pp.
II-1-II-21.
Los
Alamos
National
Laboratory,
Los
Alamos,
N.M.
VOL.
67,
1993

4036
GORELICK
ET
AL.
21.
Morellet,
N.,
N.
Jullian,
H.
De
Rocquigny,
B.
Maigret,
J.-L.
Darlix,
and
B.
P.
Roques.
1992.
Determination
of
the
structure
of
the
nucleocapsid
protein
NCp7
from
the
human
immunode-
ficiency
virus
type
1
by
1H
NMR.
EMBO
J.
11:3059-3065.
22.
Popovic,
M.,
M.
G.
Sarngadharan,
E.
Read,
and
R.
C.
Gallo.
1984.
Detection,
isolation,
and
continuous
production
of
cyto-
pathic
retroviruses
(HTLV-III)
from
patients
with
AIDS
and
pre-AIDS.
Science
224:497-500.
23.
Prats,
A.
C.,
L.
Sarih,
C.
Gabus,
S.
Litvak,
G.
Keith,
and
J.-L.
Darlix.
1988.
Small
finger
protein
of
avian
and
murine
retrovi-
ruses
has
nucleic
acid
annealing
activity
and
positions
the
replication
primer
tRNA
onto
genomic
RNA.
EMBO
J.
7:1777-
1783.
24.
Rein,
A.,
B.
I.
Gerwin,
R.
H.
Bassin,
L.
Schwarm,
and
G.
Schidlovsky.
1978.
A
replication-defective
variant
of
Moloney
murine
leukemia
virus.
I.
Biological
characterizations.
J.
Virol.
25:146-156.
25.
South,
T.
L.,
P.
R.
Blake,
R.
C.
Sowder
III,
L.
0.
Arthur,
L.
E.
Henderson,
and
M.
F.
Summers.
1990.
The
nucleocapsid
protein
isolated
from
HIV-1
particles
binds
zinc
and
forms
retroviral-
type
zinc
fingers.
Biochemistry
29:7786-7789.
26.
South,
T.
L.,
and
M.
Summers.
1990.
"Zinc
fingers"
in
metal
ion
induced
regulation
of
gene
expression,
p.
199-248.
In
L.
G.
Marzilli
and
G.
Eichhorn
(ed.),
Advances
in
inorganic
biochem-
istry
series,
vol.
8.
Elsevier
Science
Publishing,
Inc.,
New
York.
27.
South,
T.
L.,
and
M.
Summers.
1993.
Zinc-
and
sequence-
dependent
binding
to
nucleic
acids
by
the
N-terminal
zinc
finger
domain
of
the
HIV-1
nucleocapsid
protein:
structure
of
the
complex
with
the
Psi-site
analog,
d(ACGCC).
Protein
Sci.
2:3-19.
28.
Summers,
M.
F.,
L. E.
Henderson,
M.
R.
Chance,
J.
W.
Bess,
Jr.,
T.
L.
South,
P.
R.
Blake,
I.
Sagi,
G.
Perez-Alvarado,
R.
C.
Sowder
III,
D.
R.
Hare,
and
L.
0.
Arthur.
1992.
Nucleocapsid
zinc
fingers
detected
in
retroviruses:
EXAFS
studies
of
intact
viruses
and
the
solution-state
structure
of
the
nucleocapsid
protein
from
HIV-1.
Protein
Sci.
1:563-574.
29.
Summers,
M.
F.,
T.
L.
South,
B.
Kim,
and
D.
R.
Hare.
1990.
High-resolution
structure
of
an
HIV
zinc
fingerlike
domain
via
a
new
NMR-based
distance
geometry
approach.
Biochemistry
29:329-340.
30.
Towbin,
H.,
T.
Staehelin,
and
J.
Gordon.
1979.
Electrophoretic
transfer
of
protein
from
polyacrylamide
gels
to
nitrocellulose
sheets:
procedure
and
some
applications.
Proc.
Natl.
Acad.
Sci.
USA
76:4350-4354.
31.
Waters,
D.
Unpublished
results.
J.
VIROL.
